阅读:0
听报道
文 | Calderón等 编译 | 集智翻译组
导语:海鞘的蝌蚪幼体是第二个完成全部神经网络连接绘制的动物,透过这些小型的大脑,我们可以对其他物种的大脑神经网络进行进一步的探索。
在自然界,动物大脑的大小千差万别,有大到超过7千克重的鲸鱼,也有小到只有66个神经元组成的一种侏儒雄性蠕虫(Windoffer and Westheide, 1988)。然而不管是大是小,它们都有一个共同的特征,那就是组成它们的神经元都会连接形成一个复杂的网络。这种特性在其他器官是见不到的,比如肝脏,肾脏或者皮肤。因此如何绘制神经网络连接就成了神经科学领域的一个重要课题,近年来连接组学的火热也证明了这一点。
如果想要绘制神经网络的连接图谱,即展示神经元之间独特连接的连接组,则需要通过电子显微镜来观察一些超薄的神经组织切片。虽然这个方法在近些年来取得了巨大的成果,但是却只能分析相对少量的组织。举例来说,一个小鼠的脑组织大约有1cm³,是现在可以观察与分析体积量的100,000倍。这也是为什么体积在连接组学里很重要的原因。所以到目前为止,也只有秀丽隐杆线虫(Caenorhabditis elegans)这一个物种的连接组全部绘制完成。这个包含了302个神经元的连接图谱是在三十年前完成的,耗费了超过10年的努力(White et al.,1986)。
1.有史以来的“第二个”全连接图谱物种
而现在来自达尔豪斯大学(Dalhousie University)的Kerrianne Ryan, Zhiyuan Lu 以及Ian Meinertzhagen在elife杂志上报道了他们的成果:成功绘制了来自玻璃海鞘(Ciona intestinalis)蝌蚪幼虫的连接组(Ryan et al., 2016)。成年的海鞘并不怎么显眼,充满水的囊鞘在离开海水之后会自发喷射,这也是它们为什么叫做“海洋喷射器(sea squirts)”的原因(如图1A)。然而来自俄国的胚胎学家在1867年发现海鞘的蝌蚪状幼虫与脊椎动物(拥有脊椎的动物,包括从鱼到人都属于脊椎动物,Mikhailov and Gilbert,2002)有着相似的基础身体横剖结构,从而吸引了科学家们的注意。后来的分子实验进一步证明,海鞘的确是我们的“无脊椎”近亲,而现在玻璃海鞘也成为生物学领域非常重要的实验动物。
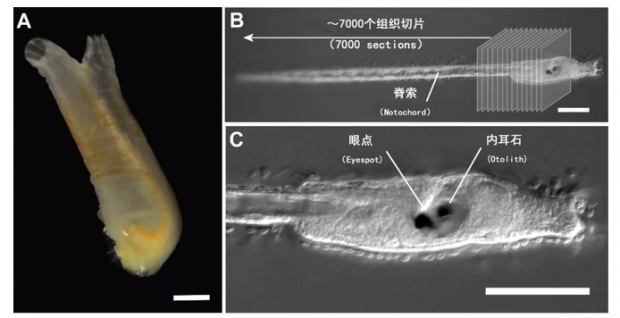
(A) 玻璃海鞘成体是固定不动的,通过滤食生存的动物。比例尺为10mm。
(B) 玻璃海鞘蝌蚪幼虫可以游动,并且有着类似脊椎动物的身体横剖面,背部有一个灵活的棒状脊索(notochord)。Ryan等人将它切分成7000个超薄切片,并用这些切片重构了海鞘幼体的连接组。
(C) 海鞘幼虫同样拥有感觉器官,包括一个在脑袋右侧的一个眼点(Eyespot)以及左侧一个可以感知重力的器官(内耳石(Otolith))。B,C图的比例尺都是100μm。
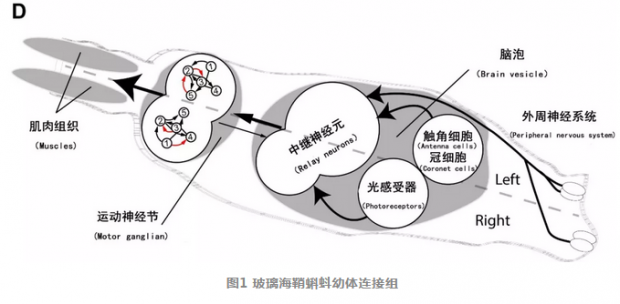
(D) 海鞘的连接组显示海鞘幼虫的神经系统包括外周和中枢神经系统,后者包括一个脑泡(Brain vesicle)以及一个运动神经节(motor ganglion)。脑泡里面又包括组成眼状斑点的光感受器(photoreceptors),冠细胞(Coronet cells)(用于感受压力),触角细胞(antenna cells)(用于感知内耳石的位置)以及中继神经元(relay neurons)。而运动神经节里面则包括直接控制肌肉收缩的神经元。从解剖结构以及神经元的连接方式来看海鞘幼虫的大脑是高度不对称的,比如说在海鞘幼虫的左边要有多得多的中继神经元,而且运动神经节左右运动神经元的连接方式也是不同的(数字1-5),红色箭头展示了左右不对称的连接方式。 照片A, B以及C 由Antonio Palladino提供。
拥有不到200个神经元的玻璃海鞘,它的神经系统已经达到了组成大脑最少所需要的极限。然而绘制和重建单样本上的神经系统的大部分神经元是一项非常巨大的工作。Ryan等人首先将海鞘幼虫切分成7000个切片,每个切片仅有十几纳米厚,并采用电子显微镜对每一个切片进行拍照。然后再对这些高分辨率的照片进行对齐,并人工地去标记神经元。最终,所有的神经元连接得到辨认和标记。通过这一工作,他们获得了玻璃海鞘幼虫几乎完整的连接图谱,这是有史以来第二个得以建立全连接图谱的物种。
2.如何理解幼虫背后的行为机制?
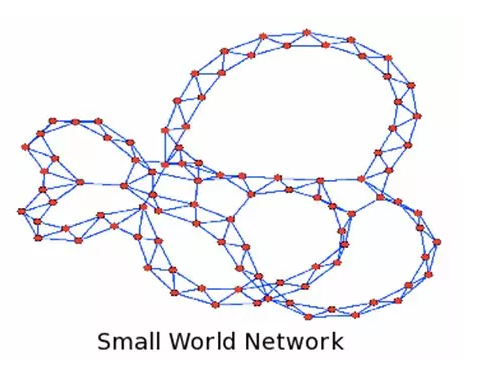
那么我们能从玻璃海鞘的连接图谱中学习到什么呢?首先这是一个被称为所谓的“小世界”网络,也就是说这个网络是由高度连接的子网络通过少量的远程连接构成的。这与秀丽隐杆线虫的连接组,社交网络以及电力网络的构成是很相似的(Watts and Strogatz, 1998)。总而言之,这177个神经元所构成的连接网络中,从一个节点到另一个节点的平均距离只需要2.7步。
而且这个连接组也表明玻璃海鞘幼虫的外周神经系统与中枢神经系统有着明显的区分。外周神经系统包括少量的感受细胞并直接与环境直接接触。中枢神经系统由几个感受系统组成,一个在头部右侧的单眼,一个可以感受重力的器官以及一些可以感受压力的细胞(如图 1C)。这些感觉系统与大脑背侧的中继细胞高度相连。这些中继神经元又与同属中央神经系统一部分的其他中间神经元以及运动神经元相连接,最终连接至玻璃海鞘幼虫的肌肉系统。
那么连接组学如何帮助我们理解海鞘幼虫的行为机制呢?首先,这类生物几乎所有行为都与它的肌肉有关系。甚至于说,在它孵化之前,非对称性的肌肉收缩也可以帮助海鞘幼虫的尾巴以每秒10次的频率摇摆并使它最终摆脱卵壳。而且它的尾巴在孵化后的持续摆动对它在游泳时改变方向也许很重要(Mast, 1921)。然而,随着海鞘幼虫年龄的增长,它行动的动力会变成依靠尾部对称性收缩来实现,而这种收缩频率可以达到每秒20-30次。此外,通过改变肌肉的行动,海鞘幼体可以对不同的刺激做出反应,诸如光线,重力以及阴影(Zega et al., 2006; McHenry and Strother, 2003),比如说海鞘幼虫会向着与重力相反的方向游动。Ryan和其同事完整绘制了负责这一相对简单行为的神经通路,从感受器到肌肉组织(如图1D)。
尽管这些行为看似很简单,但是它们并非完全独立。与此相反,这一网络允许不同层次之间的网络通信。这也提示我们尽管大脑可以整合不同的刺激信号,但是每次却只激活一种行为模式。这与我们观察到的海鞘幼虫实际的游动方式是相吻合的(Zega et al., 2006)。这些发现表明,虽然每个海鞘的幼体很小,在解剖学上看也很简单,但是构成其神经系统的神经网络是复杂而完整的。
3.打破神经系统的对称铁律?
Ryan等人还注意到玻璃海鞘连接组另一个很有意思的特征。目前大多数动物研究都表明神经网络存在着一种左右对称的规律,也就是说对于每一个左边的神经元,都有一个右边的神经元与之相对应。然而对大多数玻璃海鞘的神经系统却缺乏这种对称(如图1D)。不仅如此,即便是玻璃海鞘中一些对称性的神经元,比如说运动神经元,它们连接到其他细胞方式也是非对称的。至于为何玻璃海鞘的神经网络会表现出如此强的非对称性,我们并不清楚,但是这很有可能与海鞘流线型的身体横剖面结构有关。在所有动物的大脑中都有非对称性的身影,包括我们自己,只是所占的比例很少,因此玻璃海鞘可以为非对称性在大脑里的演化与功能研究提供线索。
海鞘的连接组图为理解大脑的工作机制提供了经验和基础。很明显,我们对这个最小的大脑的研究,还有很多工作要做。尽管只有非常少量的神经元,但距离彻底理解它还有很长的路要走。通常来说,在处理那些更为复杂的大脑之前将重心集中在这些小型大脑上是非常明智的。这一考虑也有经济上的意义,因为绘制一个小鼠的连接组的花费是做小型大脑连接组的100,000倍。或许其他从事连接组工作的研究员会被Ryan, Lu以及 Meinert zhagen的出色工作所鼓舞,进而优先考虑小型大脑的研究。
参考文献
[1]. Delsuc F, Brinkmann H, Chourrout D, Philippe H.2006. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 439:965–968. doi: 10.1038/nature04336, PMID: 16495997.
[2]. Mast SO. 1921. Reactions to light in the larvae of theascidians, Amaroucium constellatum and Amarouciumpellucidum with special reference to photicorientation. Journal of Experimental Zoology 34:148–187. doi: 10.1002/jez.1400340204.
[3]. McHenry M, Strother J. 2003. The kinematics ofphototaxis in larvae of the ascidian Aplidium constellatum. Marine Biology 142:173–184. doi: 10.1007/s00227-002-0929-z.
[4]. Mikhailov AT, Gilbert SF. 2002. From development to evolution: the re-establishment of the "Alexander Kowalevsky Medal". The International Journal of Developmental Biology 46:693–698. PMID: 12216980.
[5]. Ryan K, Lu Z, Meinertzhagen I. 2016. The CNS connectome of a tadpole larva of Ciona intestinalis (L.) highlights sidedness in the brain of a chordate sibling. eLife 5:e16962. doi: 10.7554/eLife.16962.
[6]. Tsuda M, Sakurai D, Goda M. 2003. Direct evidence for the role of pigment cells in the brain of ascidian larvae by laser ablation. Journal of Experimental Biology 206:1409–1417. doi: 10.1242/jeb.00235, PMID: 12624175.
[7]. Watts DJ, Strogatz SH. 1998. Collective dynamics of ’small-world’ networks. Nature 393:440–442. doi: 10.1038/30918, PMID: 9623998.
[8]. White JG, Southgate E, Thomson JN, Brenner S. 1986.The structure of the nervous system of the nematode Caenorhabditis elegans. Philosophical Transactions of the Royal Society B: Biological Sciences 314:1–340. doi: 10.1098/rstb.1986.0056.
[9]. Windoffer R, Westheide W. 1988. The nervous system of the male Dinophilus gyrociliatus (Polychaeta,Dinophilidae): II. Electron microscopical reconstruction of nervous anatomy and effector cells. The Journal of Comparative Neurology 272:475–488. doi: 10.1002/cne.902720403, PMID: 3417897.
[10]. Zega G, Thorndyke MC, Brown ER. 2006. Development of swimming behaviour in the larva of the ascidian Ciona intestinalis. Journal of Experimental Biology 209:3405–3412. doi: 10.1242/jeb.02421, PMID: 16916975.
编译:集智翻译组
来源:
作者:Luis A Bezares-Calderón, Gáspár Jékely
原题:Connectomics: Think small
翻译:薛伟
审校:胡鹏博
编辑:李沛欣
原文地址:
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}