
人们一直以来都坚信着达尔文“物竞天择”的进化原则。但一些物理学家认为,达尔文的进化论并不是唯一可以创造秩序的东西。对进化和生命本身的看法应该基于热力学以及这些物理定律对能量和物质流动的表述。因为,虽然生物属于一整类复杂而有序的系统,但这些系统是基于热力学第二定律存在的。
这篇发表于 2007 年的经典论文,系统阐述了热力学定律对自然选择与生命现象的解释。集智俱乐部热力学与进化论小组曾在 2009 年的读书会中研读过这篇文章,2019 年 4 月,集智第谷读书会重温经典,对这篇文章做了线上讲解分析,视频见文末。
编译:集智俱乐部翻译组
来源: PLoS Biolog
原文题目:Survival of the Likeliest?
使用热力学定律来解释自然选择和生命本身
表面上看,进化论和热力学似乎是相互矛盾的。最明显的是,热力学第二定律指出,随着时间的推移,任何系统都会倾向于达到熵的最大值,这意味着系统的秩序和可用能量均为最低水平。在一个封闭的房间里打开一瓶香水,最后香气也会消散,而有机体通常会尽量避免气味的消散,也就是所谓的死亡。反进化论者一个常见的论点就是,宇宙倾向于无序意味着自然选择不能使生物变得更加复杂。当然,也有人反对这样的观点,他们认为生物体通过一种输出熵的方式维持内部秩序并建立复杂性,即以一种形式吸收能量并以另一种更高水平的熵的形式辐射出来。Erwin Schrödinger(薛定谔)是思考这些问题最早的一批物理学家之一,他将食物描述为负熵,并认为“有机体新陈代谢的关键在于,能够成功地将自己的生命从生产无益的熵中解放出来。”[1]

达尔文的进化论并不是唯一可以创造秩序的东西。
最近,一些物理学家指出,虽然生物属于一整类复杂而有序的系统,但这些系统并不违反热力学第二定律,反而是基于定律而存在。他们认为,我们对进化和生命本身的看法同样应该基于热力学以及这些物理定律对能量和物质流动的描述。他们指出,达尔文进化论并不是唯一可以创造秩序的东西。在整个宇宙中,能量和物质的相互作用带来了规则的结构,无论是恒星,晶体,流体中的漩涡,还是天气 系统都是如此。而生物是迄今所了解的最复杂,最有序的系统,那么它们可能是同一现象的一部分吗?并且最终能通过热力学语言解释生物之间相互竞争带来自然选择过程吗?
“达尔文的自然选择进化论不是独有的过程,”圣塔菲研究所的理论物理学家Eric Smith说,“他们是基本的化学竞争性排斥的更为复杂的版本。”在去年发表的一篇论文中[2],Smith及其同事认为,自然选择是一种被称为“自组织”的高度复杂的物理过程,自组织是一种至今仍不为人知的方式,通过这种方式,能量与物质相结合生成秩序。

这种有序、自组织的系统就像专门设计出来的用于平衡能量梯度的引擎——当自组织系统持续存在时,它们产生熵的速度,比无序分子混合物产生熵的速度更快。例如,天气系统将热量从热带地区传递到极地的速度远远快于均匀的静态大气层。生命也是如此。事实上,Smith认为这可能是生命起源的原因——在地球早期的条件下,生命是释放地热能积聚的最佳方式,也是地热能释放的必然结果[3]。一旦这种生物化学过程能够进行下去,那么后续的化学和达尔文式选择(subsequent chemical and Darwinian selection )就成为了耗散掉地球上积压能量的最好途径,包括天气系统地球热物理的能量或者是太阳能。
长期以来,人们都认为,自组织系统不仅仅比无序的系统更快地平衡能量梯度,而且它的速度是所有可能中最快的一种。一些模型运用最大化熵产生原理 maximum entropy production(MEP)可以很好的预测有关地球的气候系统[4]、土星的Titan卫星[5]以及溶液中的晶体生长[6]等过程。但直到现在,MEP也只是一个假设——没有机制或理论来解释为什么系统会趋向于这种状态。而经典的热力学对此也无能为力,因为它只在封闭系统中解释熵,其中没有能量的输入输出。经典热力学没有说明像生命体这样开放的非平衡系统应该产生多少熵。
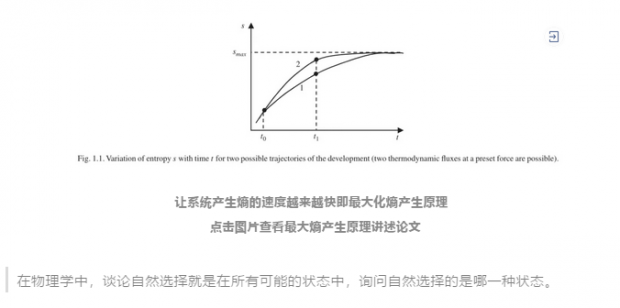
Roderick Dewar是法国农业研究机构波尔多中心的理论物理学家和生态系统建模师,他相信自己已经跨越了一些障碍。使用信息理论——可以重新制定热力学定律的数学分支,Dewar已经证明,只要系统自由“选择”其状态,不受任何强大的外力的干扰,在许多相互作用元素组成的开放的、非平衡系统中,MEP即是最可能的行为模式[7]。无论系统的微观部分在做什么,MEP的大规模状态描述了系统微观部分的无数可能中占比最多的情况。
生物学中的自然选择也能以同样的方式进行,Dewar认为:在物理学中,谈论自然选择就是在所有可能的状态中,询问自然选择的是哪一种状态。他指出,这是一个概率问题。“自然选择的状态比其他任何状态都更容易实现。虽然生物学家不这么认为,但我想假设生物学中的自然选择也是以同样的方式进行的,看看这能给我们带来什么。”
向物理系统中增加生命自然会增加产生的熵。一个充满浮游生物的池塘或一片草地吸收了更多的太阳能量,因此比无菌池或裸露的岩石产生更多的熵。地球比火星或金星更高效地将太阳光转变为微波辐射,与宇宙背景辐射更接近平衡。自然演替的生态过程,例如草原成为森林等也会增加熵的产生。在进化的过程中,生物体往往能更好地吸收能量——想想我们周围的这些物种,使用了太阳光中约40%的能量,并同时也不断释放化石燃料中的能量并将其转化为熵。这种过程能否被解释为MEP趋势的一部分,而不是一种达尔文式的留给后代的竞争?关键在于生物是否真的可以自由地达到MEP的状态,或者自然选择是否恰好是凌驾于这一过程之上的力量。

“自然选择可能不是适者生存,而是系统最可能的一种结果。”——虽然这种观点似乎很奇特,但Dewar就是这么认为的。最近,他和他的同事们用MEP理论[8]证明了ATP合成酶的结构和工作原理是可以预测的。总的来说,Dewar想要证明,从统计力学的角度可以解释最大程度摄取能量比例的生物过程,或者将化学物质从一点转移到另一点的过程。统计力学是物理学的一个领域,它解释了可预测的行为如何从大量不可预测的元素中产生。“统计理论会说,分子选择最大通量的状态,是因为这是系统中分子排列最可能的方式,”Dewar说,“也许他们选择这种状态只是因为它是最可能的状态。”与传统的进化论观点不同,这种方法允许人们对生物应该如何运作做出定量预测。“达尔文选择是一个很难量化的假设,”Dewar说,“它并没有真正给出数字。”
如今,一些生物学家也开始使用MEP。加州大学伯克利分校的生态学家John Harte说:“Dewar的证明是杰出的,对许多科学领域都有潜在的巨大影响。”生态学是可能受到其影响的领域之一,他补充道:“对理解食物网、生物体内部物质和能量分配以及气候生态系统相互作用的影响的一些初步的探索都是令人鼓舞。”
✎Box: 什么是熵?
熵是一个强大但难以捉摸的概念。这其中一个原因是几个不同的物理学分支已经能够独立地表述热力学第二定律。这意味着其他领域,如计量和生态学中,也可以使用熵的概念,于是熵在不同的系统中有不同的形式。
在热力学中,熵是无用的度量,例如温差这类能量梯度,可以用来做功。但是随着梯度逐渐变缓,能量转化为与周围环境平衡的无用的热量。在统计力学中,系统的熵是产生任何特定宏观状态的所有微观状态的可能排列的数量。最大熵是最可能的,也是最无序的状态。例如,抛1000枚硬币,最有可能,也是熵最大的状态,是500个正面和500个反面。这种形式的熵也被称为“混合度”:一杯白咖啡的分子排列数量远远大于一杯加了一层牛奶的黑咖啡。
在信息论中,熵是不确定性的度量。熵最大的系统是那些人们最不确定接下来会发生什么的系统。在非常有序的信息中,例如一串相同的字母,下一个字母是可预测的,这样的系统没有熵。而一串随机的字母非常杂乱,没有携带任何信息,且具有最大的熵。这个熵的公式是由数学家克劳德·香农(Claude Shannon)提出的,他还以自己的名字命名了一种衡量生物多样性的指数——香农指数(Shannon index)。这个指数表示了在许多类别中个体分布的均匀程度。种类越多,个体数量越均等,生物多样性就越大;这在数学上等价于熵的度量。在最多样化的生态系统中,博物学家几乎不知道她下一步会发现什么物种。
动物运动的模式就是动物们沿着地球表面进行流动的最有效率的方式
另一个物理学家试图用热力学来预测生物结构的细节信息,他就是Adrian Bejan,一位Duke大学(位于北Carolina的Durham)的工程师。Bejan没有考虑系统的微观元素,而是设计了一套他所谓的“构造法则” (Constructual law)[9],它描述了能量和物质如何在诸如河流流域这样的物理网络和诸如血管这样的生物网络中流动。Bejan的构造法则指出,对于一个流动系统,要想使它持续存在,那么随着时间的推移,它必须提供更容易流通的渠道——换句话说,它必须用更少的资源做更多的事情。在这个过程中,它使燃料使用量最小化,并使每单位燃烧的燃料所产生的熵最大化。

Bejan认为,进化是一个结构自我改造的过程,使能量和物质尽可能快速有效地流过[10] 。无论是动物交互还是河流纵横,较好的流动结构取代了较差的结构。Bejan说,这是第二支时间之箭,与热力学第二定律的混乱度相呼应。他认为,动物运动的模式,尤其随着体型的变化动物的步幅或节拍的频率的变化,使得动物在地球表面流动时能尽可能容易[11]。Bejan说:“考虑到形态的自由,流动系统将自我优化,以构建更容易的流动结构。”“动物群体在地球上运动模式遵循着与亚马逊河水在陆地上流动相同的原则。”
Dewar却不这么认为,他认为构造法则处理的是现象,而不是原因。他说:“Bejan没有解释为什么系统应该采用最优行为,而是提出它们应该这样做,然后表明这是现实。”目前还不清楚什么被最大化了——似乎是他能想到的任何东西。Bejan认为Dewar对系统最小元素的关注是不必要的:“一个人不需要进入微观来解释宏观。”
动物运动的模式是使动物尽可能容易地在地表通行。
除了物理学家对此存在异议外,许多生物学家也毫不奇怪地抵制了殖民他们学科的企图。已故的Ernst Mayr认为,生殖、自然选择和遗传等过程在物理学中没有等价性,也不能简化为物理定律,生物学应该被视为一门独立自主的科学[12]。尽管并非所有的生物先贤都这么认为: Francis Crick写道,生物学的“终极目标”应该是用化学和物理[13]来解释自己。
哈佛大学的数学生物学家Lloyd Demetrius并不排斥物理。他采用了基于统计力学的方法,把生物体当作气体中的分子来对待,并引入了一个他称之为“进化熵”的量[14]。这在数学上等同于热力学熵,但它描述的是生物体繁殖的年龄结构,而不是物理上的混乱度。在漫长的进化过程中,Demetrius认为自然选择会增加这个量,因为能够在较长时间内繁殖的生物体更善于应对有限的资源和不可预测的环境。
但在Demetrius的模型中,进化熵并没有最大化,也不会随着时间不可避免地增加。他说,热力学过程和自然选择之间存在根本的区别,生物和物理选择只在分子水平上是一体的。任何更复杂的生命系统都受到不在纯粹物理系统中运行的力量的影响。“在进化过程中,有类似于物理定律的东西,但是机制是完全不同的,”Demetrius说,“从分子到细胞和高等生物体,自然选择包括自我繁殖,而在物理学中没有自我复制的情况。这就是生命体与非生命体的区别。”
也许再过一百年,没有人会认为我们需要一套生物学理论和另一套物理学理论。
然而,对于那些被自组织系统和生命系统之间的相似之处所震撼的物理学家来说,即使是这种区别也不像看上去那么明显。但是,“生命和非生命之间存在连续统一体,两者之间的差别必须最小化。” 堪培拉澳大利亚国立大学的天文学家和天体生物学家Charles Lineweaver这样说。
Lineweaver提出了一类系统,他称之为“far from equilibrium dissipative systems”,其中包括所有维持自身能量耗散处于有序,非平衡状态的系统,例如星系、飓风以及动植物等。他认为,所有这些系统都有可能被描述为生命体,并且生命应该用热力学术语来定义。“作为一名物理学家,我正在寻找基于物理学的生命的定义,”Lineweaver说,“对此,生物学家的观点过于短浅了。”
Lineweaver还认为自我繁殖的问题实际上转移了我们的注意力。他说,认为生命必须在内部储存繁殖指令是武断的。他指出,恒星的形成依赖于前一代恒星释放元素以及改变其环境的重力。一切都取决于能源和材料所处的环境; 存储信息的地方无关紧要。Lineweaver说:“把生命的定义转变成热力学,就像达尔文说:‘嘿,我们是另一种动物’一样,它消除了生命的神秘感。”

一百年前,生物学中最激烈的争论之一是关于“活力论”的——生物是否由与无生命物质相同的化学物质构成,它们是否由生物系统特有的“生命力”构成,或者是否遵循与无生命物体相同的物理定律。一个世纪过去了,我们知道生命和无生命物体是由同样的物质构成的,并服从于相同的力量。也许再过一百年,没有人会认为我们需要一套生物学理论和另一套物理学理论来解释生命和物质系统。
“我们应该寻找共同的原则,” Dewar说,“如果存在这样的原则,我们应该能够将生物学中的自然选择与物理学中的自然选择融合在一起。动物的竞争和死亡最终即是在能源和资源的限制下发生的分子过程。”
参考文献:
[1]Schrödinger E (1992) What is life? Cambridge (United Kingdom): Cambridge University Press. 194 p.2.
[2]Hoelzer GA, Smith E, Pepper JW (2006) On the logical relationship between natural selection and selforganization. J Evol Biol 19: 1785–1794.
[3]Morowitz H, Smith E (2006) Energy flow and the organization of life. Santa Fe Working Paper 06-08-029.
[4]Paltridge GW (1975) Global dynamics and climate - a system of minimum entropy exchange. Q J Roy Meteorol Soc 101: 475–484.
[5]Lorenz RD, Lunine JI, Withers PG, McKay CP (2001) Titan, Mars and Earth: Entropy production by latitudinal heat transport. Geophys Res Lett 28: 415–418.
[6]Hill A (1990) Entropy production as the selection rule between different growth morphologies. Nature 348: 426–428.
[7]Dewar RC (2005) Maximum entropy production and the fluctuation theorem. J Phys A 38: L371–L381.
View Article Google Scholar
[8]Dewar RC, Juretic D, Zupanovic P (2006) The functional design of the rotary enzyme ATP synthase is consistent with maximum entropy production. Chem Phys Lett 430: 177–182.
[9]Bejan A (2000) Shape and structure, from engineering to nature. Cambridge (United Kingdom): Cambridge University Press. 324 p.10.
[10]Bejan A (2005) The constructal law of organization in nature: Tree-shaped flows and body size. J Exp Biol 208: 1677–1686.
[11]Bejan A, Marden JH (2006) Unifying constructal theory for scale effects in running, swimming and flying. J Exp Biol 209: 238–248.
[12]Mayr E (1996) The autonomy of biology: The position of biology among the sciences. Q Rev Biol 71: 97–106.
[13]Crick F (1966) Of molecules and men. Seattle: University of Washington Press. 120 p.14.
[14]Demetrius L (2000) Thermodynamics and evolution. J Theor Biol 206: 1–16.
翻译:SBu
审校:Krystal
编辑:林下
原文链接:

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}