阅读:0
听报道

导语
大脑,神经元构成的器官根植于许多生物体内。这是一种固态的大脑,且组成它们的元素在空间中相对固定。但是相对的,大脑是否可以拥有流动不定的存在形式?越来越多的研究发现无定形的认知网络——例如蚁群与免疫系统具有智能,它们可以被称为液态形式的大脑。
圣塔菲研究所自2017年开始一系列关于“液体”大脑与“固体”大脑主题讨论,经历了几年打磨,形成了15篇文章,于近期集中发表在英国皇家学会自然科学会报 B 上。
本文是《Liquid brains, solid brains》论文系列引言的翻译,其中预告和概述了涉及的主题,翻译的文末,我们将附上系列中其他论文的目录。
编译:集智俱乐部
来源:The Royal society
原题:Liquid brains, solid brains
为了解决信息收集存储和回应过程中产生的问题,认知网络发展一套出自成体系的解决方案。一些认知网络被描述为自适应连接网络中的静态神经元集。它们是“固态”的网络,具有明确的静态神经元集。
其他的一些智能系统由一定数量的单体组成,信息在这些单体之间交换、存储和处理。但是,这些网络没有稳定的连接和物理空间中稳定的结构。我们将这些缺乏稳定连接和静态元素的网络称为“液体”大脑,一些例子包括蚂蚁、白蚁蚁群、免疫系统、以及一些微生物和黏菌 。
固体和液体大脑之间的主要区别是什么?它们的认知潜力,解决特定问题和应对环境的能力,以及信息处理策略分别是什么? 要回答这个问题,需要将这个问题的框架搭建起来。
本文是论文系列“液体大脑,固体大脑:分布式认知架构如何处理信息”的首篇,为之序。
生命的认知结构是如何进化的
结构复杂多样的生命形式在寒武纪大爆发时期集中出现。在那时,生命的认知能力以及维持认知能力的网络使多细胞生物的存在成为可能。生命的尺度(scale)开始变得庞大,同时形成它的细胞也开始出现分工,出现有目的的运动能力以及其他协作能力。然后生命们又开始渴求更高级的感知和记忆能力。 虽然用行为适应环境的生存模式在神经元出现之前已经存在[1],但神经结构的出现和快速发展促进了新的复杂行为的诞生。
生命进化的功绩将很大程度归功于神经,它们的前体细胞,神经元细胞以及它们构成的神经回路,深刻地改变了生命存在的方式。信息,由此成为复杂生物适应周围环境的另一个重要组成部分,使生物拥有超越基因能直接赋予的更快反应能力。从寒武纪时代在生命树中出现了一条路径,描述了具有两侧对称特征的动物(bilaterians)。在这个类群内,动物的躯体具有明确定义的横向对称。如此,神经元也以一种对称的方式配置成网络。 这些神经网络最终演变成分离的神经元群,形成运行中央控制功能的系统。 第一个大脑就此形成[2],并很快成为一个强大的预测机器[3]。
运动假说提出,生物体对空间环境的积极探索是大脑进化轨迹上的一个关键节点[4 ]。 从这个角度来看,预测能力既是动物移动的原因,也是动物移动的结果。生物要想实现探索世界,它们就需要学习网络的支持[5]。 捕食可能是学习网络诞生过程中最重要的选择压力[ 6 ]。 这种强大的进化创新并不局限于神经网络,我们在这里展示出了多种相互作用的单体,它们最后演变出或同或异方式处理信息的能力。
信息处理网络可以在微生物群落,细胞内部(基因调控网络)和免疫系统中找到。 这些网络的架构和信息处理策略的多样性令人惊叹。流动的信息网络在成千上万甚至数百万只蚂蚁,或白蚁蚁群之内展开[7,8]。黏菌是一种简单的生命形式由相互连通的细胞团组成,可以解决复杂的问题。 植物似乎占据了认知网络领域的一个非常不同寻常的领域,缺乏像神经一样的结构和物理运动,但它们却进化得非常成功,并成为生态中的重要群体液体计算机和化学反应提供了一组完全不同的案例研究,其中计算和信息处理并不是明确定义的。在这种情况下,发展计划和模式形成也被看作是认知的一种形式[9]。
这些认知的例子中有什么共同之处? 是否存在基于组织结构、机制或功能的分类? 系统的物理存在形式是否是决定这个系统运算能力范围的主要限制?是否存在一设计上的原理,允许我们预测尚未被进化发现的系统?自然界的进化策略是否能够启发新形式的计算?也许合成生物学能帮助我们实现这一过程?回答这些和其他基本问题是2017年12月在圣菲研究所举办的小型研讨会的目标。会议召集了来自科学、工程领域的研究人员。他们的研究领域包括社会昆虫行为学、微生物学、合成生物学、发育和系统生物学、神经科学、计算机科学和统计物理学。几天后,参与者踏出了最初几步,建立了液态,固态脑理论。其长期目标是为认知网络的一般理论奠定基本原理。
液体大脑/固体大脑
神经元/无神经元
这个雄心勃勃的计划的出发点是对认知网络领域的高阶综述 :小型和大型的;分布式的和集中的;模块化的和分层的;生物的和人造的。 所有这些都由多个组件组成,这些组件可以交换环境和内部信号并对其做出反应来收集、存储和处理信息。
它们的非线性特性要求(在大多数情况下)超出标准定义以外的见解[10]。此外,大量单体非线性互动展示出的种群动力学很大程度取决于基本网络组件是否是能够移动的。我们确定了两个关键维度来描述不同类别的认知网络:系统的物理特性,以及神经元的存在与否。
下列四张图表示了四类认知网络,分为两个维度。分别为实际神经元是否存在和网络的物理组织是液态还是固态。
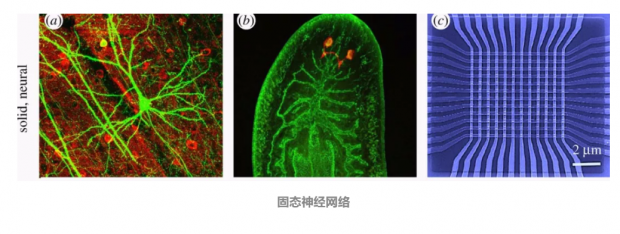
固态神经网络:
标准神经元网络(a)涉及通过分化后的,位置固定的神经细胞突触权重连接而定。较简单的生物,如涡虫(b)扁虫含有不同于图(a)中较复杂脑的神经结构。受真实神经元的启发,以硅单位生成的人工阵列(c)可以通过感知和响应环境信号来模仿其生物的一些特征,但这些刚性空间结构是没有神经元细胞的。
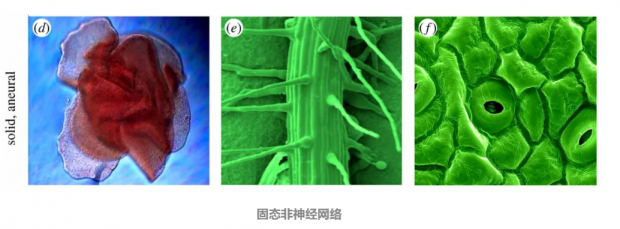
固态非神经网络:
原生动物(Placozoans)(d)完全缺乏神经元并且具有非常简单的解剖复杂性,但表现出活跃的行为。结构不同的植物也缺乏神经元,但它们的一些模块化部分,包括根(e)和叶子中的气孔(f)都具有一定自动能力,属于“固体”子类。

液态神经网络:
液体大脑由有神经系统或神经样组分的组分形成,例如(g)蚂蚁或(h)白蚁的蚁群及其在机器人领域的人工对应物(i)。在这些液体大脑中,每个成分都有自己的固体大脑。
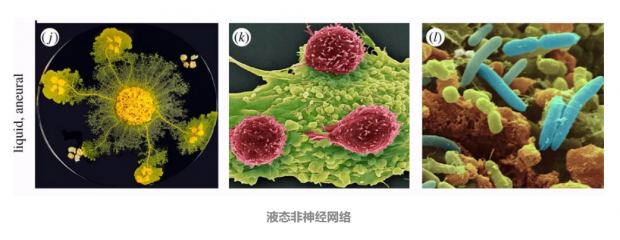
液态非神经网络:
第二类主要的液体脑包括缺乏内部大脑的运动个体,例如(j)黏菌(Physarum),(k)免疫网络和(l)微生物群落。它们中没有类似神经的组分,但在许多方面,这些系统解决了复杂问题,表现出学习和记忆,并根据环境条件做出决策。最后,有证据表明免疫系统和微生物群落在某种程度上与宿主生物的大脑存在相互作用。
第二个维度立即涉及了如何定义“大脑”的问题,这个问题在神经科学教科书中很少得到解答(存在例外,见[11])。标准的描述是,事实上大脑是一个由(通常大量)神经元组成的器官,其功能基于通过环境相互作用改变连接模式。 连接模式由突触比重(synaptic weights)所描述,随时间迁移,个体与环境互动增加而决定。早期讨论大脑的性质以及如何定义它们的研究[12]只考虑脊椎动物的大脑。 Pagan [ 13 ]和Turner [14]将问题扩展到更广泛的情况,研究范围从最小的大脑到迥然不同(比如固体和液体)的认知系统。
过去几十年中学界对大脑的研究蓬勃发展,导致了脑科学研究中,以网络为出发点的研究已经成为核心[ 15 ]。 连接体这一概念(connectome)涉及解剖和功能神经元网络上不同级别的复杂性,目前是在神经科学中的主流意见。 原因则在多个尺度上显而易见。 通过突触末端连接的单个细胞的小神经回路已经成为大多数经典学习和控制方法背后的关键因素。
分布式智能:细菌虽小但不愚蠢
甚至在复杂的神经元网络进化之前,微生物就发现了可以应对环境压力的群体结构。因此,生存与合作密切相关,合作需要集体内部新颖的交流方式。引用James Shapiro的话说:“细菌虽小但不愚蠢”[16]。 在微生物尺度的一个广为人知的集群行为是群体感应(QS)。这个能力使得细细菌能协同工作[17]。 QS允许细菌来监测群体范围内其它细菌的存在,在某些情况下导致出现群体级协调反应。 这个现象论证了微生物菌落做出群体决策的能力。另一方面,生物膜的集体行为被细菌群落中长距离的电信号通讯证明。 [ 18 ]。 Martinez-Corral 等[19]则研究皮质脑活动和生物膜动力学中可能存在的类似化学信号 。
黏菌(Physarum polycephalum)是一种引人注意的生物,一个通过单个细胞聚合来揭示集体行为的例子。虽然这种有机体是单细胞的(但它们存在多个细胞核),聚集起来的黏菌们在探索周围环境时显示出高度复杂的空间形态模式。 黏菌以连贯的方式移动和扩散细胞质的节律脉冲。 这种脉动活动发生在连接的等离子体管组成的复杂网络中。丰富的化学和生物电成分形成驱动力,遍布在黏菌的整个行为反应过程中,允许网络根据外部和内部的信号做出安排和调整。黏菌体内的这些管道可在低,高养分条件下经历变薄或增厚,而科学家已经可以将它们用作能够解决优化任务的生物计算机,从找到短路径甚至解决迷宫到计算几何问题皆可。 在所有的这些情况下,环境空间结构,排斥因素和吸引因素的浓度等信息相对应的“输出”被编码在黏菌的形态学特征中。

黏菌也可以通过使用外部记忆来探索身边的环境[21],它们被观察到做出与营养需求相关的最佳决策[22]。 正如[23]中所讨论的那样,它们也表现出对负面持续刺激的适应性,即一种共同的适应性反应(一般由具有神经的生物表现出这种反应)。 这一发现支持了这样一种观点,即没有大脑的系统也可以在适当的条件下从经验中学习,从而区分不同的信息来源。
认知世界的边界可以通过考虑最简单的“固体”大脑并了解它们如何完成工作来描述,并将之与类似简单的液体实例相比[24]。 涡虫(Planarians/ flatworms)是第一个使用大脑生物的候选人[2,13]。 特别有趣的是,它发育和认知现象的紧密联系。正如[25]中所指出的那样 ,信息处理功能在固体大脑出现之前就已经诞生很久了。涡虫可以再生他们的每一部分身体[26],实验研究表明,记忆在它们的头部缺失之后仍然会存在(见 Shomrat & Levin [27]及其中的参考文献)。 这些结果表明基于神经的现象与体细胞记忆之间存在深层联系。重要的是,涡虫对外界的许多响应可以映射到一个吸引子图与形态的对应关系。 导致这些基于吸引子响应的动力学可以在不同类型的非神经硬件中实现,尽管我们仍然缺乏用于描述这些系统的通用理论框架,正如[ 9 ]中所讨论的 。
固体的认知区域在自然界中有不同程度的跨群体共享 ,例如跨越组织、生活习惯、与不同的生命周期。这种现象特别存在于植物这种个体极端的案例中[28,29]。 早在达尔文的专著中就已经认识到植物的认知潜力[30]。在那里他指出了植物对外部信号和环境线索所表现出的有趣反应。植物可以表现出有趣的计算反应[28]。并且在18年12月 ,“植物智能”[31]的概念也得到了发展(虽然存在一定程度的争议)。植物中跨越尺度的通信,从叶子中的气孔网络到通过根系发送的信号的能力特别引起了人们的注意。 这些例子表明人类仍需要更好地理解植物中的信号处理[28],内容包括且不限于遗传开关,模拟计算机,种子散播和发芽 [32]。 特别需要注意,植物种子阶段的过程涉及植物生命周期中“能够移动”的时间段。
上述例子展现了认知分布式系统广阔世界的一瞥,这些系统可以描述为各种非线性互作的大量单体组成的复杂网络[33]。 超越这些网络所扮演的特定功能之上的,这些网络肯存在一些基本的,共通的属性。近三十年前人们就提出了一些网络潜在的共性[34],包括大多数连接模型共有的两个关键属性:
(1)任何时间节点下,任何变量之间的联系都是有数量极限的。
(2)节点之间的连接是可变的,因此网络的强度和/或连通性模式都是可以随着时间变化而变化的。
Farmer讨论(Farmer's discussion)[34]指出了这些共同点,它可以作为网络科学统一的“罗塞塔石碑(Rosetta Stone*)”(见[35])。 在液体大脑的背景下,寻找关键属性的想法特别有用。与吸引子有关的动力学现象在液体大脑的案例中是常见的(说不定是普世的),暗示在研究液体大脑中可以使用统计物理学方法[36,37]。我们需要一些新型模型,让这些模型足以包括几个具体或实际的研究案例。 这些案例可能不一定基于生命系统,化学计算机也能展示丰富的计算性[38]。 我们在这里没有严格定义什么是液体,而是可能会用一系列数学方法描述集体运动,去解释运动实体的规律。(见[39],其中的参考文献) 。
注:罗塞塔石碑是一块古埃及多语言石碑,古埃及文字的破译很大程度依赖于它,引申为基石。
除了统计物理学之外,信息和计算也是问题的核心概念,我们缺乏完整的模型来描述哪些计算问题可以通过液体大脑很好地解决,哪些不能[40]。 这部分的总结在下图(a\b\c\d)中展示,什么样的形式方法(formal approach)是可取的往往并不清楚。 Wood [41] 的建议指出,要了解不同类型大脑的能力范围需要在它们进行的信息处理和计算之间进行更正式的分类,计算与信息相关的术语在现阶段仍然可以互换使用。 其他强大的计算范式, 如水库计算(Reservoir Computing)在处理生物信息时可能更合适[42]。

蚁群(a,由Guy Therauiaz提供)寻找最优路径的过程可以理解为最少作用量问题(least-action problem),其中通过优先选择具有最高信息素浓度的路径来发现最短路径。

黏菌,Physarum polycephalum(b)是单细胞生物,也使用最小动作逻辑来解决几何和图论的问题,其中包括找到迷宫的最短路径。

目前人们尚不清楚如何对涡虫(Planarians)(c)能解决的问题进行分类,这是一类扁形虫,其神经系统是双侧对称结构组织的,具有固体的“大脑”和两只眼睛。涡虫以过去事件为输入进行分布式记忆(与人类类中心化的记忆相对):它们的形态可通过生物电信号重新编辑,这可能在认知和发展中起重要作用。
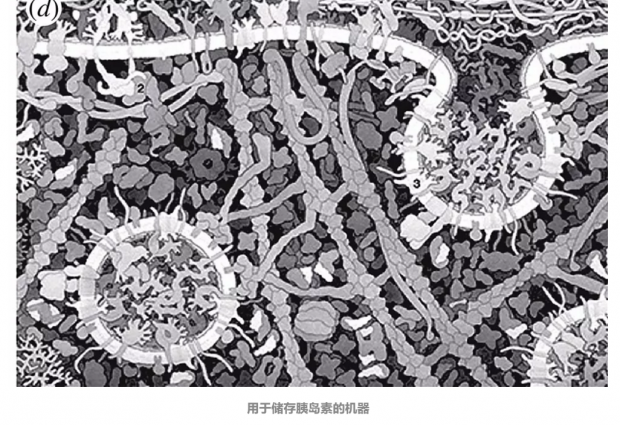
最后,单细胞包含多种信息处理现象,包括复杂的阶梯级联,遗传-代谢相互作用和泡状结构相关的计算,例如(d)中显示的用于储存胰岛素的机器。
Vining 等[43]开发了液体细胞自动机,来证明液体系统如何在没有复杂结构的情况下进行计算 。网络的移动性被证明可以增加移动单元之间的信息流,这些单元会随着时间的推移遇到其他单元且与之产生联系(Calovi 等, [44])。这表明液体系统,比如是白蚁,就能使用环境的物理结构作为计算的重要输入。Kao等[45]描述了一种不同类型的混合系统,其中动物种群运动过程中存在的模块化结构意味着即使在更大的液体系统中,某些模式的交互也能更持久和稳固(就像在固体大脑中),并且这可以导致在复杂环境中更好的集体决策。
在蚁群和黏菌的网络中,我们可以找到使用最少作用量原理(least-action principle)解决问题的类似思路。 计算的执行是黏菌结构形成(pattern-forming)的结果。但是,认识到人类实际上为黏菌生成最短路径的行为提供了一种人造的边界条件,实验的这些条件对我们理解这些智能系统极为重要。人为提供的条件多大程度定义了这个单细胞系统能执行的计算的丰富程度? 在涡虫中,如上所述,记忆的存储并非中心化的,这也展示了一种完全不同形式的再生相关计算(regeneration-related computation)。
在较小的尺度中,复杂的计算是由细胞内部的化学网络承担的,在这里我们发现了丰富的可能性。
从类似布尔型开关,到有限自动机,最后到膜计算机。 所有这些替代方案都在非常嘈杂的条件下运行。虽然这个级别的计算模型通常用基因调控网络来描述,但它们也发生在不同的网络上[46-48](Silva-Rocha,等2011).所有这些生物类型的计算的整合将需要合适的方程来描述“计算形态空间(computational morphospace)” [49] 。
让我们回到这次的主题讨论——液体大脑与固体大脑。这些主题共同形成了一张关于认知网络整合观点发展过程的地图。这对了解液体与固体系统的相对计算优势有着重要的科研意义。在未来,这些理解可能会发展出重要的,新颖的配置传统计算的方式。随着计算继续从人类的桌面上转移到整个物理世界中,我们预计以分布式,移动和自治组件为特征的液体计算将变得比现在更加重要。
参考文献
1. Jékely G. 2010Origin and early evolution of neural circuits for the control of ciliary locomotion. Proc. R. Soc. B 278, 914-922. (doi:10.1098/rspb.2010.2027) Link,ISI, Google Scholar
2. Pagán OR. 2014The first brain. The neuroscience of planarians.Oxford, UK: Oxford University Press. Google Scholar
3. Llinas RR. 2001I of the vortex: from neurons to self. Cambridge, MA: MIT Press. Crossref, Google Scholar
4. Llinás R. 1987‘Mindedness’ as a functional state of the brain. InMindwaves (eds C Blakemore, SA Greenfield), pp. 339–358. Oxford, UK: Basil Blackwell. Google Scholar
5. Miguel-Tomé S. 2018The influence of computational traits on the natural selection of the nervous system. Nat. Comput. 17, 403-425. (doi:10.1007/s11047-017-9619-0) Crossref, ISI, Google Scholar
6. Monk T, Paulin MG.2014Predation and the origin of neurones. Brain Behav. Evol. 84, 246-261. (doi:10.1159/000368177)Crossref, PubMed, ISI, Google Scholar
7. Gordon DM. 2010Ant encounters. Princeton, NJ:Princeton University Press.Crossref, Google Scholar
8. Wilson EO. 2012The social conquest of Earth. New York, NY, USA: W.W. Norton & Co. Google Scholar
9. Manicka S, Levin M. 2019The Cognitive Lens: a primer on conceptual tools for analysing information processing in developmental and regenerative morphogenesis. Phil. Trans. R. Soc. B 374, 20180369. (doi:10.1098/rstb.2018.0369) Link, Google Scholar
10. Forrest S. 1990Emergent computation: self-organizing, collective, and cooperative phenomena in natural and artificial computing networks.Physica D 42, 1-11. (doi:10.1016/0167-2789(90)90063-U) Crossref,ISI, Google Scholar
11. DeSalle R, Tattersall I. 2012The brain: big bangs, behaviors, and beliefs. New Haven, CT: Yale University Press. Google Scholar
12. Netsky MG. 1986What is a brain, and who said so?Br. Med. J. Clin. Res. Edn 293, 1670. (doi:10.1136/bmj.293.6562.1670)Crossref, Google Scholar
13. Pagán OR. 2019The brain: a concept in flux. Phil. Trans. R. Soc. B 374, 20180383. (doi:10.1098/rstb.2018.0383) Link, Google Scholar
14. Turner JS. 2019Homeostasis as a fundamental principle for a coherent theory of brains. Phil. Trans. R. Soc. B 374, 20180373. (doi:10.1098/rstb.2018.0373)Link, Google Scholar
15. Sporns O. 2010Networks of the brain. Cambridge, MA: MIT Press.Crossref, Google Scholar
16. Shapiro JA. 2007Bacteria are small but not stupid: cognition, natural genetic engineering and socio-bacteriology. Stud. Hist. Philos. Sci. C: Stud. Hist. Philos. Biol. Biomed. Sci. 38, 807-819. (doi:10.1016/j.shpsc.2007.09.010)Crossref, Google Scholar
17. Waters CM, Bassler BL.2005Quorum sensing: cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 21, 319-346. (doi:10.1146/annurev.cellbio.21.012704.131001) Crossref, PubMed,ISI, Google Scholar
18. Prindle A, Liu J, Asally M, Ly S, Garcia-Ojalvo J, Süel GM.2015Ion channels enable electrical communication in bacterial communities. Nature527, 59. (doi:10.1038/nature15709)Crossref, PubMed, ISI, Google Scholar
19. Martinez-Corral R, Liu J, Prindle A, Süel GM, Garcia-Ojalvo J.2019Metabolic basis of brain-like electrical signalling in bacterial communities. Phil. Trans. R. Soc. B374, 20180382. (doi:10.1098/rstb.2018.0382) Link, Google Scholar
20. Adamatzky A (ed.) 2016Advances in Physarum machines: sensing and computing with slime : Springer. (doi:10.1007/978-3-319-26662-6) Crossref, Google Scholar
21. Reid CR, Latty T, Dussutour A, Beekman M. 2012Slime mold uses an externalized spatial ‘memory’ to navigate in complex environments. Proc. Natl Acad. Sci. USA 109, 17 490-17 494. (doi:10.1073/pnas.1215037109)Crossref, ISI, Google Scholar
22. Dussutour A, Latty T, Beekman M, Simpson SJ. 2010Amoeboid organism solves complex nutritional challenges. Proc. Natl Acad. Sci. USA 107, 4607-4611. (doi:10.1073/pnas.0912198107)Crossref, PubMed, ISI, Google Scholar
23. Boussard A, Delescluse J, Pérez-Escudero A, Dussutour A.2019Memory inception and preservation in slime moulds: the quest for a common mechanism.Phil. Trans. R. Soc. B 374, 20180368. (doi:10.1098/rstb.2018.0368) Google Scholar
24. Vining W, Esponda F, Moses M, Forrest S. 2019How does mobility help distributed systems compute?Phil. Trans. R. Soc. B374, 20180375. (doi:10.1098/rstb.2018.0375) Link, Google Scholar
25. Baluska F, Levin M. 2016On having no head: cognition throughout biological systems.Front. Psychol. 7, 902. (doi:10.3389/fpsyg.2016.00902) Crossref,PubMed, ISI, Google Scholar
26. Gentile L, Cebria F, Bartscherer K.2011The planarian flatworm: an in vivo model for stem cell biology and nervous system regeneration.Dis. Models Mech. 4, 12-19. (doi:10.1242/dmm.006692) Crossref,PubMed, ISI, Google Scholar
27. Shomrat T, Levin M. 2013An automated training paradigm reveals long-term memory in planarians and its persistence through head regeneration. J. Exp. Biol. 216, 3799-3810. (doi:10.1242/jeb.087809) Crossref,PubMed, ISI, Google Scholar
28. Duran-Nebreda S, Bassel GW.2019Plant behaviour in response to the environment: information processing in the solid state. Phil. Trans. R. Soc. B 374, 20180370. (doi:10.1098/rstb.2018.0370)Google Scholar
29. Oborny B. 2019The plant body as a network of semi-autonomous agents: a review. Phil. Trans. R. Soc. B 374, 20180371. (doi:10.1098/rstb.2018.0371) Link, Google Scholar
30. Darwin C. 1875The movements and habits of climbing , UK: John Murray. Google Scholar
31. Trewavas A. 2014Plant behaviour and intelligence. Oxford, UK:Oxford University Press. (doi:10.1093/acprof:oso/9780199539543.001.0001) Crossref, Google Scholar
32. Bassel GWet al. 2011Genome–wide network model capturing seed germination reveals coordinated regulation of plant cellular phase transitions. Proc. Natl Acad. Sci. 108, 9709-9714. (doi:10.1073/pnas.1100958108)Crossref, ISI, Google Scholar
33. Camazine S, Deneubourg JL, Franks NR, Sneyd J, Bonabeau E, Theraulaz G. 2003Self-organization in biological systems.Princeton, NJ: Princeton University Press. Google Scholar
34. Farmer JD. 1990A Rosetta stone for connectionism. Physica D 42, 153-187. (doi:10.1016/0167-2789(90)90072-W) Crossref, Google Scholar
35. Farmer JD, Packard NH, Perelson AS. 1986The immune system, adaptation, and machine learning.Physica D Nonlin. Phenom. 22, 187-204. Crossref, Google Scholar
36.Solé RV. 2011Phase transitions.Princeton, NJ, USA: Princeton University Press. Crossref, Google Scholar
37. Piñero J, Solé R. 2019Statistical physics of liquid brains. Phil. Trans. R. Soc. B 374, 20180376. (doi:10.1098/rstb.2018.0376)Link, Google Scholar
38. Adamatzky A. 2019A brief history of liquid computers. Phil. Trans. R. Soc. B 374, 20180372. (doi:10.1098/rstb.2018.0372) Link, Google Scholar
39. Vicsek T, Zafeiris A.2012Collective motion. Phys. Rep. 517, 71-140. (doi:10.1016/j.physrep.2012.03.004) Crossref,ISI, Google Scholar
40. Mitchell M. 2011What is computation?Ubiquity 2011, 1-7. (doi:10.1145/1940721.1944826)Google Scholar
41. Wood CC. 2019The computational stance in biology.Phil. Trans. R. Soc. B 374, 20180380. (doi:10.1098/rstb.2018.0380) Link, Google Scholar
42. Seoane LF. 2019Evolutionary aspects of reservoir computing.Phil. Trans. R. Soc. B 374, 20180377. (doi:10.1098/rstb.2018.0377) Link, Google Scholar
43. Vining WF, Esponda F, Moses ME, Forrest S. 2019How does mobility help distributed systems compute?Phil. Trans. R. Soc. B374, 20180375. (doi:10.1098/rstb.2018.0375) Link, Google Scholar
44. Calovi DS, Bardunias P, Carey N, Turner JS, Nagpal R, Werfel J.2019Surface curvature guides early construction activity in mound-building termites. Phil. Trans. R. Soc. B 374, 20180374. (doi:10.1098/rstb.2018.0374)Link, Google Scholar
45. Kao AB, Couzin ID. 2019Modular structure within groups causes information loss but can improve decision accuracy. Phil. Trans. R. Soc. B 374, 20180378. (doi:10.1098/rstb.2018.0378) Link, Google Scholar
46.Bray D. 1990Intracellular signalling as a parallel distributed process. J. Theor. Biol. 143, 215-231. (doi:10.1016/S0022-5193(05)80268-1) Crossref, PubMed,ISI, Google Scholar
47. Benenson Y. 2012Biomolecular computing systems: principles, progress and potential. Nat. Rev. Genet. 13, 455-468. (doi:10.1038/nrg3197) Crossref, PubMed,ISI, Google Scholar
48. Silva-Rocha R, Tamames J, dos Santos VM, de Lorenzo V.2011The logicome of environmental bacteria: merging catabolic and regulatory events with Boolean formalisms. Environ. Microbiol. 13, 2389-2402. (doi:10.1111/j.1462-2920.2011.02455.x)Crossref, ISI, Google Scholar
49. Solé RV, Macia J. 2013Expanding the landscape of biological computation with synthetic multicellular consortia. Nat.
圣塔菲“液态大脑和固态大脑”论文专辑
Introduction: Liquid brains, solid brains
引言:液体大脑,固体大脑
Ricard Solé, Melanie Moses, and Stephanie Forrest
链接:
The Cognitive Lens: a primer on conceptual tools for analysing information processing in developmental and regenerative morphogenesis
认知透镜:形态发生过程中的发展与再生过程中的基本信息分析概念工具
Santosh Manicka and Michael Levin
链接:
The brain: a concept in flux
大脑:一个流动的概念
Oné R. Pagán
链接:
Homeostasis as a fundamental principle for a coherent theory of brains
自稳态在大脑连贯性理论中是基本原则
J. Scott Turner
链接:
Metabolic basis of brain-like electrical signalling in bacterial communities
类脑电信号在微生物群落中传播的代谢基础。
Rosa Martinez-Corral, Jintao Liu, Arthur Prindle, Gürol M. Süel, and Jordi Garcia-Ojalvo
链接:
Memory inception and preservation in slime moulds: the quest for a common mechanism
记忆在黏菌中的激发与保存:关于共同机制的探讨。
A. Boussard, J. Delescluse, A. Pérez-Escudero, and A. Dussutour
链接:
Plant behaviour in response to the environment: information processing in the solid state
植物的环境适应行为:固态条件下的信息处理。
Salva Duran-Nebreda and George W. Bassel
链接:
The plant body as a network of semi-autonomous agents: a review
植物体作为半自动单元网络:一个综述。
Beata Oborny
链接:
Statistical physics of liquid brains
液态大脑的统计物理基础。
Jordi Piñero and Ricard Solé
链接:
A brief history of liquid computers
液体计算机简史
Andrew Adamatzky
链接:
The computational stance in biology
C. C. Wood
链接:
生物中的“计算立场”
*延伸自哲学名词"意向立场(Intentional Stance)"意味一种评价生物中计算概念质量的手段。
How does mobility help distributed systems compute?
移动性如何帮助分布式系统进行计算?
William F. Vining, Fernando Esponda, Melanie E. Moses, and Stephanie Forrest
链接:
Surface curvature guides early construction activity in mound-building termites
表面曲率能指导白蚁早期的巢穴搭建。
Daniel S. Calovi, Paul Bardunias, Nicole Carey, J. Scott Turner, Radhika Nagpal, and Justin Werfel
链接:
Evolutionary aspects of reservoir computing
水库计算技术的进化视角。
Luís F. Seoane
链接:
Modular structure within groups causes information loss but can improve decision accuracy
群体中的模块化结构造成信息丢失,同时提高决策精度。
Albert B. Kao and Iain D. Couzin
链接:
翻译:陈曦
编辑:陈安林
原文地址:
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}