阅读:0
听报道

2021年诺贝尔奖颁发给乔治·帕里西 (Giorgio Parisi) 以表彰他对复杂系统理论的开创性贡献,特别是“发现了从原子到行星尺度的物理系统中无序和波动的相互作用”。除了原子和宇宙,帕里西还专门研究过鸟群的波动。故事源之于有一天他在罗马的火车站上空看到了成千上万只鸟儿成群结队地飞翔,鸟群没有统一的指挥却能如同一个整体自由变换形状,这一现象让他着迷。帕里西于是派出了一组物理学家,他们拍摄记录了大量的鸟群飞行数据。基于此,他们用统计物理方法计算分析了数十万只鸟如何形成一个整体。
对一个鸟群而言,最智能的状态应该是某种“团结紧张”的状态。这种状态恰好处在“有序”与“无序”之间,类似于相变的临界点处,此时,一个群体既能保持其稳定性,又能保证个体的信息在群体中有效地传递。
撰文 | 傅渥成

图1. 鱼群的集体运动
最早尝试研究动物集体运动的人是日本的鱼类学家,他们早在六七十年代就提出了鱼类集体运动中的基本原则,然而因为太过于超前,他们又只用日语发文章,这些研究并没有得到重视。而到了八十年代,随着计算机科技的发展,动物集体运动的原则重新被计算机科学家 Reynolds 发现了,这一工作发表在计算机图形学的会议上。我们今天见到的许多动画、电影中的大场面,通常都是用类似的方法让计算机生成的。在这篇文章中,Reynolds 总结了群体运动模拟的几个核心的要点:
体积排斥:每个模拟的个体(boid)都会占据一定的体积,而这些体积永远不会相交,即每个个体都会避免与最近的个体发生碰撞。
速度对齐:每个个体与其近邻的个体们保持速度同步。
聚集倾向:粒子不会倾向于独立行动(粒子群保持维持),每个个体会尽量与附近个体靠近,避免被孤立。
有了这些基本要点,我们就确定了集体运动中个体之间的相互作用,当许许多多的个体聚集在一起时,动物的群体就可以产生各种各样复杂的运动模式。我们已经知道,在鸟群中,相邻的鸟的速度会趋向于平行,但仅仅知道这些还并不够,我们想知道这种相邻的鸟与鸟的相互作用究竟会在多大的范围内影响鸟群的运动。事实上,虽然每个鸟只受到附近较少的几个近邻的影响,但这种影响竟然可以遍及整个群体!

图2.2021年诺贝尔物理学奖三位得主。一半奖金授予了Giorgio Parisi,表彰其对复杂系统的开创性研究
2021年诺贝尔奖得主 Parisi 曾经与 Cavagna 等人合作,对鸟群的运动进行了长期的观察,他们用两个摄像机拍摄同一个鸟群,从而重建出鸟运动的三维坐标(这与我们在电影院观看 3D 电影的基本原理是相同的),然后从这样一个 3D 的影片中算出来各个时刻,群体中的各只鸟分别处在怎样的位置,又根据相邻各帧画面,计算出鸟的速度,从这样的视频中我们可以得到很多有用的信息,这里我们介绍其中最有趣的“临界”特征。
论文题目:
Scale-free correlations in starling flocks
论文地址:
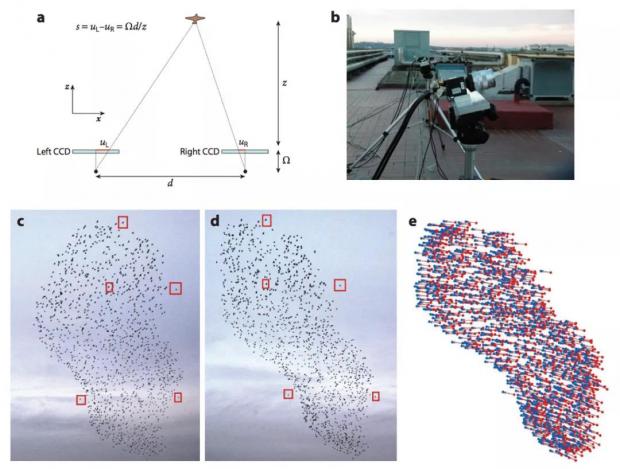
图3. 实验装置及原理图
一只鸟因为各种不确定性因素、外界刺激或者突发奇想,稍稍改变了它的运动方向,那么它的这种行为能影响到多大的范围呢?这里的“影响力范围”就是“关联长度”。分析关联,对于物理问题的分析非常重要,文小刚教授曾经在他的《量子多体理论》中提到:“我们可以测量的其实只是关联函数。我们不禁很想用关联函数来定义世界上的物理理论,关联函数可能就代表着我们世界的真实。”在舍恩伯格的《大数据时代》中,“关联”(相关)跟“因果”是相对的,在物理学里,直接的相互作用就对应于“因果”,例如两只相邻的鸟为了防止相互碰撞而产生相互排斥的效果,如果不是“因为”鸟 A 如此靠近,鸟 B 也不至于要到改变运动方向的“结果”;而“关联”则更多的是因为间接的相互作用所造成的,例如处在鸟群的外围、直接观察到了捕食者的鸟跟位于鸟群另一侧的鸟之间显然不存在直接的相互作用,但它们的运动依然能相互影响,这即为“关联”。这种关联性就可以用“关联函数”来描述。
怎样用关联函数描述鸟群中不同个体之间运动情况的关联呢?尽管整个鸟群在朝着某一个共同方向以平均速度 v 运动,但鸟群中的诸多个体可能自己的运动速度会与这个平均速度的方向或大小有所偏离。我们可以计算鸟的速度偏差之间的关联,即考虑在鸟群中挑出距离为 r 的两只鸟(i, j),让它们各自真实的运动速度分别减去平均速度,得到相对速度,然后计算相对速度的内积,再然后,把所有这样的距离为 r 的鸟都选出来计算内积,这一内积的平均值即为 C(r)。
如图 4 中(a)(b)所示,距离很近的两个鸟当然更倾向于同进退(关联接近 1,不然就会撞在一起),而距离远到一定的程度,C(r) 的值会降低到 0,在鸟群的运动问题中我们把 C(r) = 0 时 r 的取值 ξ 定义为“关联长度”,而当 r>ξ 时,鸟与鸟的速度关联可能为负,即这些鸟倾向于往相反的方向运动。如果还对速度的大小和方向的涨落进行进一步的区分,可以得到如图 4 所示的结果。如图 4 的(c)(d)中所示,对于不同大小的鸟群,其关联长度与鸟群的尺寸成正比,因而这种关联可以遍及整个鸟群。在临界态,对于无限大的系统,关联长度甚至可以趋向于无穷。
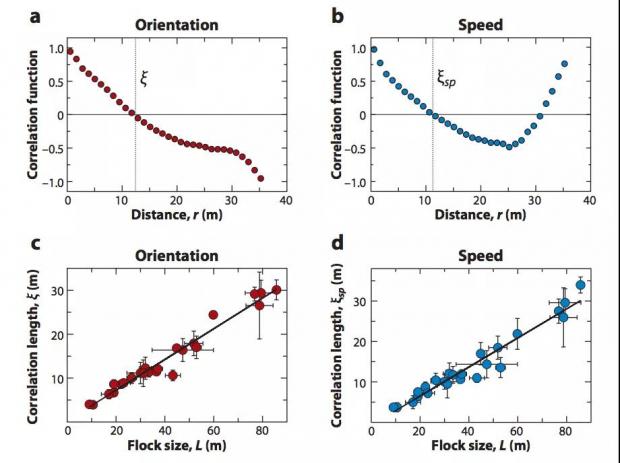
图4. 鸟群运动的(速度涨落)关联函数。这里的图 (a)和(c)对应的是速度方向的关联,图 (b)和(d)对应的是速度大小的关联。而图(a)(b)计算的是关联函数的距离依赖关系,图(c)(d)计算的是关联长度随着鸟群大小的变化关系,从图中可以看到关联长度与鸟群的大小是成正比的。
直观地看,关联函数可以理解成一种”影响力”,在一个鸟群中,平均地来看,较长的关联长度意味着各个鸟都有较大的影响力范围。如果鸟群在运动中没有达到临界,那么对于不同大小的鸟群而言,关联长度会更像是一个常数,这看起来是自然的,因为在描述集体运动的各类计算模型中,每个鸟的速度只与跟它们距离小于一定值的鸟的速度存在”相互作用”。在没有到达”临界”的情况下,在一个有 100 只鸟的鸟群中,一只鸟运动方向的改变,大概会影响与这只鸟距离小于 1 米范围内的其它同类,如果是更大规模的鸟群,如 1000 只鸟的群体,一只鸟的“影响力”范围仍然会是大约 1 米。然而如果鸟群达到了”临界”,让人感觉意外的事实发生了,正如我们在各种物理系统的临界态中所看到的那样,这种“影响力”就可能会变得非常长。
“影响力”变长并不是一件简单的事情。不同的体系中可能出现非常类似的“长程关联”现象,但这些长程关联出现的原因可能并不相同。在有的体系中,因为存在“领导人”的角色,而所有个体都无比尊崇领导人的一切决定,因此个体间就出现了这种长程关联。而有的体系或许因为一些组织形式使它们产生了这种长程关联,例如 Ising 模型所描述的磁体当其位于相变临界点附近时,这种长程关联就会存在,其关联长度会趋向于无穷。不管产生长程关联的原因如何,从观测和统计的角度来看,在系统中出现了某种与系统本身尺寸无关的关联性,这种长程关联也被叫做“无标度的关联”(scale-free correlation)。处在临界态的系统普遍具有这种无标度的关联。虽然每个鸟只受到附近较少的几个近邻的影响(保持速度的一致性),但在群体中任意一只鸟的速度如果发生变化(这种变化可能是由于发现了障碍物或者天敌所造成的),那么这种速度的变化不只会对这只鸟的邻居们产生影响,而且可以遍及整个群体。这种无标度的长程关联正是鸟群“敏感性”的来源。
有意思的是,但在各种不同的生物系统中,这种长程关联竟然也普遍存在。为什么鸟群倾向于在临界点附近工作呢?如果一个鸟群过于团结(类似于有序的固体),那么它们将会盲目地朝着一个方向飞,此时,如果有一个个体注意到前方的障碍物,在其它个体没有看到障碍物的情况下,他将没有办法阻碍集体的力量,最终,整个群体将撞上障碍物;而如果一个鸟群过于松散(类似于无序的气体),那么它们就没有办法组织起来完成那些个体无法实现的行为(如一起对抗天敌)。对一个鸟群而言,最智能的状态应该是某种“团结紧张”的状态。这种状态恰好处在“有序”与“无序”之间,类似于相变的临界点处,此时,一个群体既能保持其稳定性,又能保证个体的信息在群体中有效地传递。
临界的特征对生物群体来说有着重要的意义,在草原上迁徙的羊群,天空中飞翔的鸟群,河流中洄游的鱼群都常常会遇到相似的问题,因为捕食者可能会从各个方向靠近来捕猎群体中的某些个体,甚至有的时候捕食者也会展开群体的攻势——这些情况就像自习的教室外有老师走过一样危险。这时候,这些动物们会需要快速做出反应,于是我们看到鸟群在空中不断变换着形状,羊群突然聚集在一起,或者鱼群组合成一条看起来更大的鱼的样子,这种灵活应变的能力体现了生物的某种集体智能,这也是简单的有序或者无序态难以实现的。
大脑的工作原理与鸟群也有相似之处。在大脑中,信息的有效整合需要有“长程关联”的存在。近年来,随着各种实验技术的发展,我们可以用功能性核磁共振(fMRI)等实验直接测量大脑皮层中不同区域之间神经信号发放之间的关联,这种关联也是长程的,并且大脑皮层中距离距离较远的区域的放电情况还可能出现反关联,这些性质表明,大脑也是处在临界态的。这种特性帮助大脑始终保持在最佳的工作状态——稳定性与可塑性的最佳平衡。幸运的是,我们的大脑在稳定性和可塑性之间达成了一种巧妙的平衡,我们的大脑恰好处在一个最具适应性的临界点上。
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}