按:《生命之流》系张江老师2008年至2009年,利用闲暇时间所构思的一本科普集。主要内容包括:从复杂到生命、自主生命体简史、重新发现时间、从永动机到人工智能、连接上帝的纽带——熵等章节。现将已公开部分刊出,希望能够和大家共同学习。
1 从复杂到生命
“今天你复杂了吗?”,这是一位朋友为了讽刺当今科学研究中的随波逐流行为的一句调侃。因为时下复杂系统已经成为了一个科学研究中的热门领域,似乎不管是什么领域的研究都要跟复杂、网络扯上关系,因为你不复杂就不时髦。然而,这门新兴学科除了能给我们兜售一些诸如幂律、网络、涌现等概念之外,还会给我们带来什么样的深层次启发呢?它究竟要回答和解决一类什么样的问题?它将引领我们去向何方?这些问题使我陷入了深深的思考……
春天到了,我漫步在小河边。地上的小草刚刚发芽,几只刚刚苏醒的小虫在那里快速爬行,划出了一系列复杂随机的曲线。一阵暖洋洋的春风迎面扑来,一只灰色的小虫摇摇晃晃地沿着复杂的曲线爬到了我的脚下。然而,当我定睛一看才知道,那并不是一只虫子,而是一块普普通通的石头子儿。石子由于风的作用,在崎岖的路面上颠簸形成了复杂的运动曲线,才会勿让我把它看作是“活”的小虫。
这一情景让我陷入了沉思,究竟什么是“活”这个属性呢?如果仅从行为上判断,石子和小虫可能都会让我们觉得它们是活的,因为它们的行为同样的复杂。但我们却知道石子是死的,因为它的行为复杂性并非来源于它的内部,而是来源于外力风的作用和复杂的地形。小虫却有本质的不同,它们能够按照它们“自我的意识”来决定将朝哪个方向去觅食。
所以,我们很清楚这个世界存在着两类完全不同的东西,一类被我们称为活的,它们能够按照“自我”的意识,主动地进行运动;另一类则是死的,它们永远是被环境和物理规则驱动的被动运动着。然而,这两种事物之间的分界线真的如此清晰吗?要知道,即使是活的小虫也是由一群分子构成的,这些分子无非也遵循着物理规则被动运动。因为,科学在19世纪就已经证明,生命体不存在所谓“活素”(vitalism)的物质以及那个特殊而神秘的“活力”(vital force)。那么,究竟小虫子那种主动的活性从哪里来呢?
有一部3D动画叫作The Inner Life of a Cell 。

通过这个形象而精彩的动画,我们可以窥探到一个细胞内部的分子世界,这哪里是细胞啊,它简直就是一个人类社会!微管是一条条高速公路,一些蛋白质分子就像载货的汽车一样运输着营养物质(图1.1中的那个分子装置),线粒体是一个大的发电厂、细胞核中的DNA相当于人类社会中的中央首脑在发号施令。更有意思的是,这群分子有条不紊的复杂运动却能导致细胞整体呈现出活性来!当我们观察细胞整体的行为时,它明显地活了,因为它可以按照“它自己”的意识游向葡萄糖浓度高的地方去捕食,也可以按“它自己”的意识避开有害化学物质。一种只有生物才有的高层次的主动性体现出来了。
按照科学的解释,活细胞的这种主动性是蕴藏在成千上万个分子的物理、化学相互作用之中的整体涌现属性!然而,这种属性究竟是怎样涌现而出的呢?要知道,更详细的微观物理、化学知识并不能给这个问题的回答提供太大的帮助。比如图1.1所示的那个分子的运动完全可以从微观上用物理化学定律解释清楚,但是为什么这个小家伙要沿着微管朝这个方向而不是另一个方向运输物质呢?显然,最好的解释是说,因为细胞整体在这个方向缺少营养物质,所以细胞会给这个小家伙发送信号,使得它朝这个方向运输物质。
可是,请不要忘记,我们这个整体细胞恰恰就是由这成千上万个分子组成的呀,那么是哪一个具体分子能够代表细胞的利益发号施令呢?答案是不存在这样的分子!细胞就是细胞,它的整体是一个独立的个体!我们不能把这个整体还原成一个具体的分子!
其实细胞和分子的这种关系与我们争论已久的人类社会中个人同集体的关系非常相近。试想,某国家被外敌侵略,需要征集本国志愿兵。从个体上看,没有哪个人愿意参兵,因为这对每个人都是不利的,他们需要冒生命危险。然而,国家这个整体却需要有这群冒死的士兵,因为他们的牺牲能够保护国家大多数人的利益,虽然这个大多数人具体是哪一个人并不清楚。
所以,国家是发号施令的独立个体,它能按照“它自己”的意愿来命令它下面的个体来完成抵抗外敌的任务,尽管这个独立个体恰恰是由微观的个人构成的。我们说人类国家也是一个活生生的有机体!
没错,这个问题已经逐渐从具体的学科知识中慢慢浮出水面,而成为一个复杂系统的问题了。实际上,我们就是要问:“究竟什么样的系统才是有生命的活系统?”。与薛定谔那个著名的“什么是生命”的问题略有不同,在这里,我们已经把生命看作是一个系统的属性,而不仅仅是物理属性。所谓的系统属性就好像我们的软件一样具备某种对硬件的“超越性”。
举个例子来说明这种软件对硬件的“超越性”,我们知道“生命游戏”是一个简单的计算机程序,我们可以用屏幕上的黑、白方格按照规则来演绎该游戏;同样,如果我们把一幢大楼看作一个元胞空间,大楼窗户中的灯光的开关看作黑、白两种细胞状态的话,那么只要这些窗户开灯关灯也遵循生命游戏的规则,那么整个游戏的动态就可以在整幢大楼上重演!(现在很多大楼都装上了这样的大楼霓虹灯)。
更有意思的是,“滑翔机”这个小家伙也会在整幢大楼上遨游,因为它完全是一种独立于底层硬件的软件Pattern。按照这个类比,“生命”也是一种具备“超越性”的系统属性。它可以用一堆分子作为硬件来实现,也完全可能用一群人类个体作为硬件来实现。这样,遥远宇宙太空中很有可能存在着某种由星体相互运动构成的超级生命个体。当然,这一切的前提就是我们能够找到那个系统的“活性”出来,即究竟“生命”是一种什么样的软件?
实际上,这个问题恰恰就是现代复杂系统中的核心问题。系统科学发展到20世纪70年代以后人们才开始真正面对复杂系统。传统的控制论、系统论感兴趣的是一类由开关、机械装置构成的死系统,因为这类系统的因果规律相对清晰。例如一艘巨型轮船,尽管它的部件设计非常复杂,但是它总是需要人来操纵、指挥,这样轮船的运动完全可以归结为舵手的指挥,一切结果都可以归结为简单的原因,所以系统不具备活性。然而,对于细胞、社会、互联网这样的系统来说,似乎不存在一个控制者在那里(或者我们可以说那个控制者就是系统自己)。因果变得复杂,系统开始呈现出“活系统”才具备的特性。这种“活性”在具体的环境中就会演绎出一系列具体的问题。例如,社会系统的活性就体现了国家整体利益和个体成员利益的冲突。同样,对分子的运输行为的解释也必须从整体细胞的角度出发,因为细胞才是一个活个体。
有了这个“更大的问题”,我们不再困惑。所有的具体研究都将在这块更大的拼图中找到自己的位置。而第一步就是要解决:“究竟生命是一种什么样的软件?”。为了找到这个问题的答案,我不得不花费一本书的篇幅来讨论它。
2 生命自主体简史
上一章,我们指出来“生命”是一种存在于系统之上的软件。而复杂性科学的首要任务就是要破译这种软件。为了把我们的讨论更加形象化,我画出下面的图来表示一个系统。
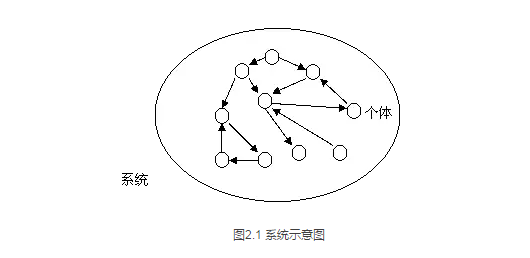
如图2.1所示,系统就是一个多个个体(小圆圈)通过各种相互作用(小箭头)组成的整体。实际上,这里的小箭头既可以表示相互作用又可以表示相互关系。因此,不严格地说,这些小的圆圈就可以看作是系统的硬件,而小箭头排列方式就是系统的软件。不同的小箭头连接方法,以及作用方式就导致了系统整体属性的千变万化。因此我们说生命是一种可能的软件结构其实就是说有生命的系统的相互连接方式具有某种特定的模式。那么,这个特定的模式是什么呢?
开始认真思考这个问题的不是财大气粗的美国人、欧洲人,也不是以擅长整体思维方式自居的中国人,而是两名来自南美的不起眼的智利人:Humberto Maturana和Francisco Varela。20世纪50年代,Maturana是通过思考“每个人的眼睛看到的可能都不是同一种红色”为起点来思考生命本质这个问题的。也就是说,他认识到,要解释生命这个现象,我们必须“钻到”生命里面去。到了1972年,他和Varela合作出版了《Autopoiesis and Cognition: The Realization of the Living》一书,为生命的系统解释Autopoiesis(自我创生)理论奠定了基础。
首先,对于细胞生命来说,系统的个体就是一个个分子,而Maturana和Varela关注的个体之间的关系是一类称之为生产的关系。我们知道,在细胞内部,每一秒钟都发生着成千上万个化学反应。无疑,每一个具体的化学反应都是非常不同的,但是我们总可以把这些反应概括成一种由若干元素生产若干元素的生产关系,例如:
A+B+C-->B+D+E
A与B和C混合生成B, D,E(这里B又可以看作是该反应的一种催化剂)。因此,无论化学反应如何复杂,它们总可以看成是节点和节点之间的生产关系。进一步,活细胞中的这种生物化学生产关系构成的网络具备什么样的特性呢?Maturana和Varela回答说,细胞内的生产网络应该是一个自我闭合的整体。
自我闭合的特性可以解释为,系统中的任意一个元素都必须是由该系统中的其他元素生产的。因此下图中的(a)和(c)都是自我闭合的生产网络,而(b)就不是。原因是(b)中的节点C并不是由系统中的其他元素(A或者B)生产的。
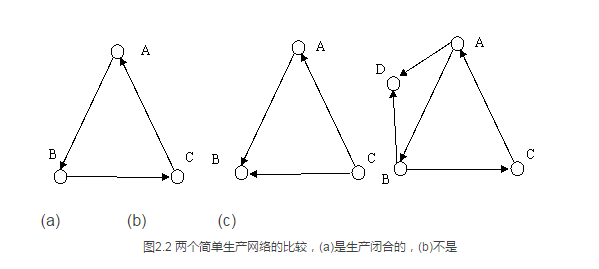
我们不难想象,图2.2(a)和(c)中的网络一旦存在就会一直稳定存在下去。原因是系统中的元素可以相互生成。假如我们人为地删除(c)中的D节点,那么它又会被A或者B重新生成。进一步如果图(a)中的节点C被删除了,虽然短时间内A分子得不到生产,但只要有一定量的B存在,B就会再创造出新的C来,而C又会继续创造A,从而使得原先的网络得以恢复。这恰恰解释了生命有机体中广泛存在的自我修复现象。当我们的手指划破了一个小口子,经过一段时间,这个小口子就会自己修复、闭合上。自生产网络就给自我修复现象提供了理论基础。
然而,Maturana和Varela的自创生理论没有仅仅停留在生产闭合网络的阶段上。他们认为图2.1所表示的系统还忽略了非常重要的东西,这就是系统所在的环境。
因此,要想完整的表达出这个关系,我们必须修改图2.1为图2.3所示:
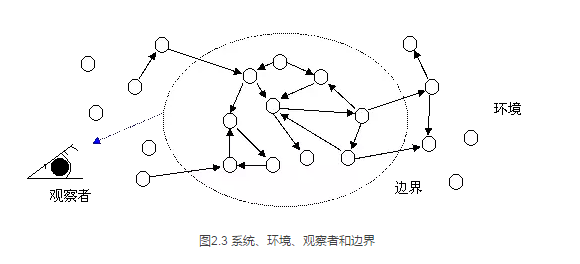
原则上讲,任何一个系统都是和它的环境通过相互作用连接成一个整体的。系统和环境之间的边界并不是先天就客观存在的。所以,如图2.3,我们所说的系统和环境之间的边界无非是外界的观察者对系统进行区分的结果,即边界并不存在。讲到这里,你会质疑到:不对,真实的细胞是有明确的边界的,这是一个双层的磷脂分子构成的细胞膜。难道这层膜是观察者观测出的结果吗?当然不是,Maturana和Varela所说的边界更多的是指区分这一概念。试想一下,假如你的尺度非常小,小到了原子的量级,那么我们看到的无非是一群模模糊糊的电子云,这时,那层清晰的细胞边界还存在吗?原来的细胞还存在吗?这就好比我们看不到国与国之间的清晰边界一样(在现实中,国界线都是一片连续的自然景观),细胞和环境的边界也取决于观察者所处的观察尺度。
因此,按照Maturana和Varela的观点,边界甚至是系统本身都是一个观察者所做出的区分结果。换句话说,对于细胞内部的分子来说,并不存在着所谓的边界、系统和环境。每个分子只知道跟自己相邻的分子发生着相互作用。而这些相邻分子有可能属于细胞之内,也很有可能属于细胞之外。因此对于细胞分子来说,即使是那层磷脂双层膜也不过是一连串的分子罢了。只有当我们的观察尺度跳到整个细胞的层面,我们才能明显地感觉到那个边界的存在,那个整体的活生生的细胞也才存在。所以,假如这个世界没有观察者的话,世界的本原就应是一片混沌的分子汤。然而,什么是那个观察者呢?观察者一定是我们人类吗? Maturana和Varela的回答是肯定的。然而,我却认为这个事情没这么简单,这个问题我们后面会进一步详细讨论。
通过综合前面讨论过的自生产和边界这两个条件,Maturana 和Varela就给自创生系统(Autopoietic system)下了这样一个定义:
自创生系统就是一个动态的系统,它是一个由个体生成的网络整体,该整体满足:
通过相互作用反复生产产生它们自己的网络;
用一个空间上的可观察整体来实现这个网络。该整体要能产生出与它所在的环境相分离的边界。
因此,自创生理论明确地指出来,生命这个软件就是:一个具备可观察边界的自我生产闭合系统。注意,他们的理论并没有限定实现这两个条件的具体硬件是什么,这也就意味着自创生系统可以是人类社会、计算机程序等其他的东西。正是因为如此,著名的社会学家Niklas Luhmann才得以把Autopoiesis理论抽象出来应用于社会学系统论的研究之中(请参考Luhmann的有关著作,这里不做详细讨论)。
到此,似乎Autopoiesis理论已经对上一章的问题,“什么样的系统是有生命的活系统”给出了很好的解答。然而,当我们尝试用Autopoiesis理论去解释那个自主觅食的小虫的时候却发现我们并没有比以前进步多少。为什么单细胞小虫子会朝向食物游去?Autopoiesis理论会告诉你这必然是因为细胞内部的分子们发生着复杂的相互作用从而推动了细胞向某个方向运动,这导致了一个高层次的可观测行为就是:细胞主动觅食了。显然要解释生命的主动运动性,仅仅具备自我生产的特性和可观察边界是不够的。换句话说,相比较生命软件来说,Autopoietic软件的版本太低了,它还不足以支持系统高级别的操作,诸如自主运动。
开始正儿八经地考虑小虫自主运动的系统性问题的人是大名鼎鼎的生物学家Stuart Kauffman,他因为写作《宇宙为家》而闻名。也是在上个世纪70年代左右,Kauffman为了研究生命起源问题提出了一个与自创生系统非常相似的自催化网络模型,如图2.4所示。
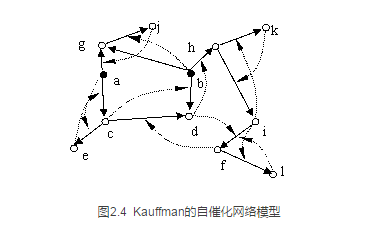
图中每个节点都是一种化学物质,实边是化学反应箭头,虚边是催化关系。例如该图中的càd的箭头就表示c可以反应生成d,同时需要催化剂f来催化。系统中的每个节点(除了黑色的食物节点以外)都是由同一个网络中的其他节点催化并反应生成的,因此它是催化闭合的。
与Maturana和Varela的自我生产闭合网络相似,Kauffman的自催化网络也是这样一个自我闭合的网络,只不过这个网络中多了一种叫作催化反应的边。同时,这个自催化网络模型也比自生产网络更加贴近现实生物体了,因为该网络考虑到了食物的情况。在图2.4中,黑色的节点a,b就是两种食物化学物质,它们并不从系统内部生成,而是必须由外界环境供给。但是,在自生产网络中,Maturana和Varela认为如果外界环境充满了细胞所必须的食物资源,那么系统对食物的要求就可以被暂时忽略。尤其是对于观察者来说,他们可以近似把一个忽略掉食物的闭合自生产网络看作一个整体(如图2.3)。
然而,Maturana和Varela所忽略的东西不仅仅是食物分子,还包括了另外一种完全不同的东西:流动!在Kauffman的模型中,这种由化学反应所构成的流动是明显的。例如从a出发,沿着网络的实箭头,我们能够明显看出三条流动路径,a-->g-->j, a-->c-->e和a-->c-->d。也就是说一旦a分子在环境中出现,那么系统就会通过化学反应创造出g物质或者c物质,进一步g物质又可以生成j,c又可以生成e或者d。一个由化学反应构成的因果关系沿着化学反应网络流动传播开了。
我们知道,很多化学反应在没有催化剂的作用下会很缓慢地发生,一旦有了催化剂,反应速度会提高上百倍。因此,一个自催化的网络系统不仅能够像自生产系统一样把自己维持住,而且它还具备一种使得内部反应流加速的现象。因此,a,b一旦提供给该网络,整个系统就会因为自催化作用而自加速起来,从而为自身提供更高的活性。
进一步,因为生物体中有些化学反应是可以释放能量的,所以,系统整体就可以由这些化学反应获得对外界做功的动力,这就为生命整体的自主运动提供了前提条件。因此,在《科学新领域的探索》(《Investigation》)一书中,Kauffman明确地给自主主体下了一个定义:
自主主体就是一个生物化学网络,该网络具备下面两个条件:
网络整体应该是自催化的;
该网络应能够完成至少一个卡诺循环
相比较Autopoietic系统的定义,Kauffman忽略掉了可观察边界的条件,而加上了卡诺循化条件。所谓卡诺循环就是指系统吸收能量、释放能量对外做功,并返回到状态原点的过程。汽车上的发动机就是在不停进行着卡诺循环。
很清楚,Kauffman的自主体模型对于我们理解自主生命又进了一步,它一方面可以反应系统内部流动的自加速现象,另一方面也指出从能量角度看,自主运动必须要能实现卡诺循环过程。这进一步给我们指示出我们要前进的方向:时间、速度、流动和能量。

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号