阅读:0
听报道

导语
一项计量研究揭示了代谢网络惊人的灵活性,为进化中的拓展适应(exaptation)提供了新的证据。
编译:集智俱乐部翻译组
来源:Quantamazine
原题:Evolution as Opportunist
进化中充斥着机会主义。被病毒感染过的宿主受益于疾病媒介留下的遗传物质;代谢酶通过眼睛的晶状体折射光线;因为颅骨间隙,哺乳动物的婴儿更容易穿过产道;更典型的,现代鸟类的祖先在登上天空之前,羽毛就出现在化石中。
这些例子表明,进化会根据需要征用现有性状特征,来实现新的用途,而这又表明性状的当前功能可能并不是最初的进化原因。
拓展适应
1982年,Stephen Jay Gould和 Elisabeth Vrba将这个现象命名为『拓展适应(exaptation)』。如字面意思所示,拓展适应是相对于适应(adaptation)的概念——适应通过自然选择来实现当下功能,而拓展适应则是为了新用途征用现有性状。例如,陆地动物用肢体行走是一种拓展适应,因为肢体骨骼的排列顺序最初是为了控水而形成;与之相对的,骨骼形状和肌肉组织的变化则是适应。
自提出以来,拓展适应这个概念一直饱受争议,主要因为在进化的历史语境中,很难区分拓展适应与适应所发挥的作用。征用性状的证据仍仅限于某些案例,例如羽毛的演变。不过在形态学、行为学和分子领域中,拓展适应日益增多的证据,让一些生物学家怀疑,这种现象在进化中的作用可能比常识认为的更大。
量化拓展适应
Nature的一项新研究首次(可能)尝试了全面识别了潜在的拓展适应。在新陈代谢系统中,该研究首次量化了拓展适应的作用。
论文题目:
A latent capacity for evolutionary innovation through exaptation in metabolic systems
论文地址:
苏黎世大学的Andreas Wagner和Aditya Barve通过计算机建模创建了随机的代谢系统,该系统需要消耗一种能源(fuel),通常也可以消耗从未使用过的其他能源。因此,如果一个假想的生物体被剥夺了通常的食物来源,它也可以接受全新的能源。在这种情况下,转换能源的能力导致了拓展适应。

分子进化生物学家 Joe Thornton 说:“我认为,越来越清楚的是:拓展适应在生物重要过程的进化中非常重要,现在有越来越多的证据印证了Gould和Vrba的发现。”
实验探究:拓展性适应隐藏的潜能
识别拓展适应需要回顾进化历史,这点很难在大多数生物学性状上实现。通过模拟进化、测试结果,Wagner 和 Barve 转换思路,回避了这个问题,而专注于新陈代谢,用计算机模拟代谢反应(reaction)的网络。生物体通过代谢反应来分解食物、产生生存和成长所必需的分子。
他们想知道:如果通过人工选择让网络使用特定碳源,如葡萄糖,它还可以使用像腺苷和乙酸盐这样的其他碳源吗?
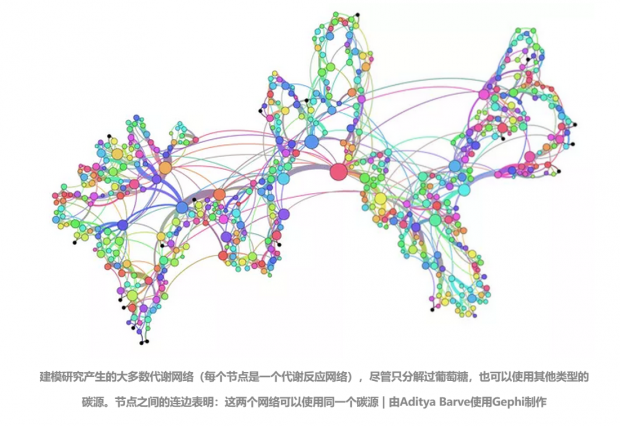
因为这一广度的研究不能用真实的有机体,最开始,Barve和Wagner采用了细菌大肠杆菌所使用的1,397-反应网络的模型。他们试图通过交换大肠杆菌网络的代谢反应和随机选择的已知代谢反应,推演进化网络。虽然自然界的众多代谢反应尚未被完全研究清楚,但新陈代谢相对容易理解,而且比其他系统更普遍、更易于控制。
研究者为上述代谢反应的交换设定了要求:不论代谢反应如何交换交换,代谢网络必须始终能够利用葡萄糖。这个设定起到了自然选择的作用,过滤了那些会引起功能失调的代谢反应交换。
Barve和Wagner制造了500个代谢网络,每个都交换了5,000次。然后他们测试网络,能否代谢除葡萄糖之外的49种其他碳源中的任何一种。事实证明,96%的网络可以使用多个碳源,平均可以使用将近5个碳源,见下图b。
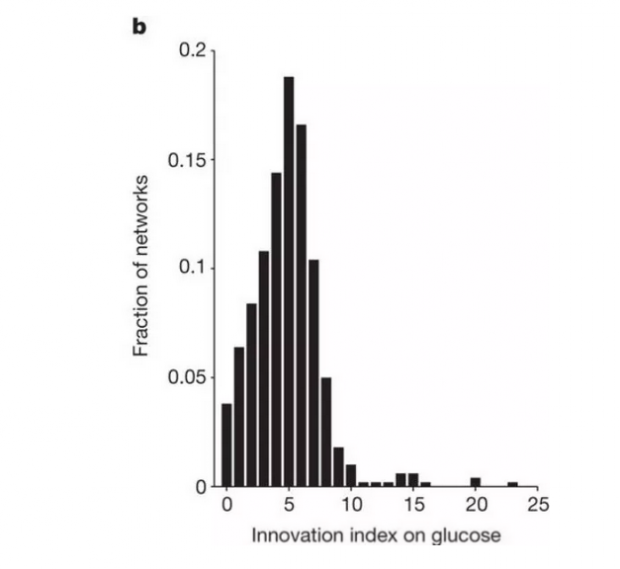
换言之,一种适应(葡萄糖)伴随着多种潜在的拓展适应(利用其他碳源)。
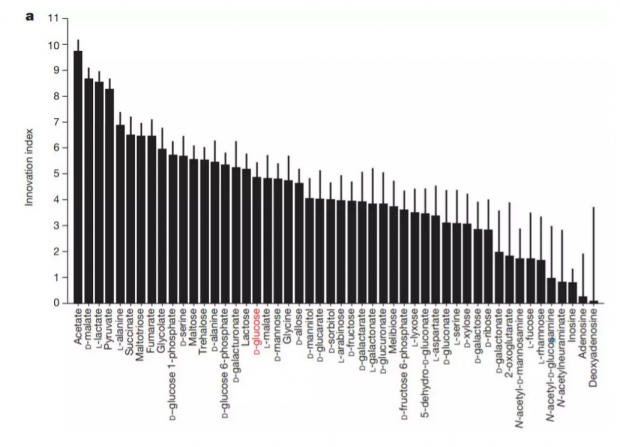
结果不仅限于葡萄糖驱动的网络。Wagner和Barve重复实验,选择使用其他49种碳源分子,发现大部分随机创建的网络可以在多种碳源上发挥作用,见下图a。
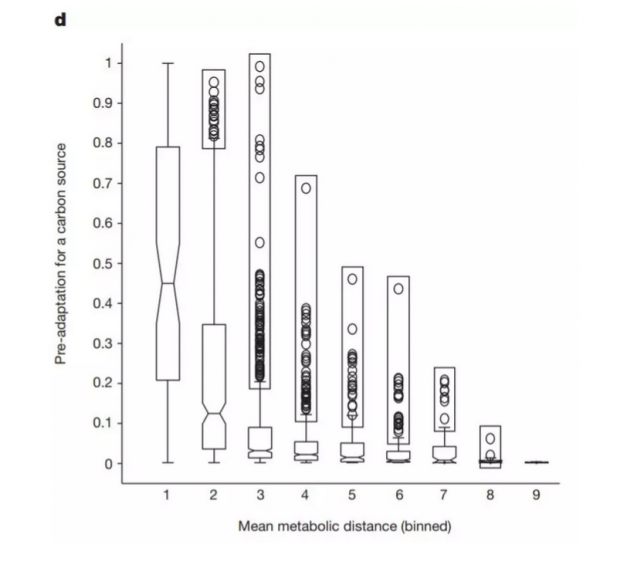
他们还发现,这种灵活性无法用所谓的“碳源之间的代谢接近(metabolic proximity)”来解释。下图b、c展现了不同的平均代谢距离(mean metabolic distance),按照该理论,网络应该更容易接受图b中代谢路径较短的新碳源。
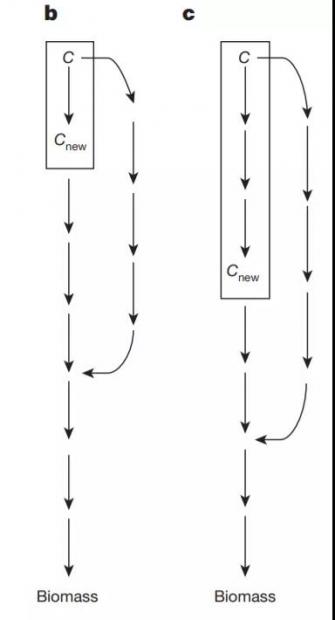
但是实验表明,葡萄糖驱动的网络没有表现出该倾向,见下图d。Wagner表示:“如果代谢接近是拓展适应的唯一解释,那就没意思了。拓展适应可能是生物化学产生作用的必然结果。”
相反,网络的复杂性似乎决定了它的灵活性;网络中的反应越多,发生拓展适应的可能性就越大。Wagner说:“许多有机体的确可以设计成更简单的结构,研究结果表明,进化中的复杂性可能蕴含着重要的潜在优势。”
新陈代谢之外
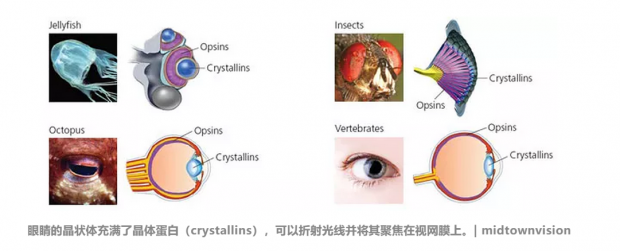
眼睛的晶状体充满了晶体蛋白(crystallins),可以折射光线并将其聚焦在视网膜上。晶体蛋白似乎是从其他无关的地方借来的。美国国立卫生研究院的名誉科学家Joram Piatigorsky指出,例如,在心脏和其他地方都发现了αB-晶体蛋白,保护其他蛋白质免于过大压力。其他晶体蛋白也可以催化代谢反应。
Barve和Wagner的工作促进了更多分子层次的实验。例如,Thornton研究了激素及其受体的进化,它们就像锁和钥匙一样匹配。他认为,在适当的环境中,可以征用配对的一半,产生新的激素受体系统。
31年前,Gould和Vrba认为,病毒带来的重复的DNA序列,称为“转座子”(transposons),起初可能没有直接作用,但以后可能会带来很大的优势。从那时起,研究表明,转座子在妊娠的进化中起着重要作用。耶鲁大学的进化生物学家,Andreas Wagner的前博士导师Günter Wagner教授表示,转座子来自病毒,但是可以发挥病毒之外的功能。初始原因和实际功能没有直接关系。
扭转天平:适应vs扩展适应
对新陈代谢的研究表明,新性状有积极意义,因为它能引发拓展适应。事实上,适应和拓展适应的比例相差很大。例如,人工选择使网络可以依靠葡萄糖生存,该网络平均可以利用将近5个其他碳源。Barve和Wagner认为,这促使人们反思对有益性状起源(origins of beneficial traits)的假设。

想象一个情景,一位微生物学家发现隔离的新细菌可以利用相当常见的碳源维生。那么这位微生物学家会说,这是一种适应,细菌过去为了生存发展出了这种性状,因此可以利用该种碳源维生。但是观察结果表明,这不一定全对,也许这只是一种衍生性状。
Wagner说:“如果我们发现的拓展适应普遍成立,那么将很难区分适应性性状和非适应性性状。”
即使在本研究之前,也很难区分这两个概念:适应与拓展适应,即非适应性性状或适应其他目的的性状。Gould和Vrba承认,二者有相互作用,而且,任何复杂特征都是两者共同作用的结果。
然而,其他人则认为,区分适应与扩展适应是不可能的,Gould和Vrba对扩展适应的定义是多余的。英国杜伦大学(Durham University)的进化生物学家Greger Larson表示:“没有任何东西是为了现在的功能而设计的。”他和他的同事们发现,在进化生物学文献中,“拓展适应”相对于“适应”的使用频率逐年下降,因为二者缺乏明确的划分——他们建议重新定义这个词。
因为过去选择压力的模糊性,很难说任何特质具有真正的适应性。鸟类和蝙蝠的翅膀可以被认为是手臂的拓展适应;然而,随后的结构改变不能称为适应,美国自然历史博物馆的脊椎动物古生物学家Mark Norell解释说,因为我们谈论的是历史事件,这是没法测试的。
但是,有些人反驳说,拓展适应和适应确实是两个截然不同而有意义的现象,尽管这种区别可能比较微妙。Thornton在一封电子邮件中写道:“事实上,几乎现存的一切都修改自以前的形式,但这并不是决定性的。”他和其他人所说的决定性因素是指自然选择。

Thornton提供了两个例子:如果新的突变使酶能够去除环境中的农药污染,那么解毒作用就是一种适应,也就是说,它是自然选择的结果。另一方面,如果负责调节一个过程的激素被征用于调节第二个过程,那就是一种拓展适应,因为激素不是通过自然选择进化来调节第二个过程。
Barve和Wagner理论方法的优势在于:他们绝对可以证明拓展适应的潜力而不受任何历史背景的约束。通过随机生成代谢网络,他们能够回避真实微生物会有的进化包袱。但要真正评估进化中拓展适应的作用,他们需要在验证在生物体中验证结果。这是他们接下来的目标。
尽管问题很难,Wagner表示研究组仍在努力解决中。
翻译:杨清怡
审校:李周园
编辑:王怡蔺
地址:
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}