阅读:0
听报道
引言
意识重要,还是潜意识重要?这个问题还可以分解为:有为重要,还是无为重要?招式重要,还是内功重要?剑宗厉害,还是气宗厉害?… 等等一系列重要衍生问题。
现在,一篇论文已经对这个基本问题进行了正面的回答:潜意识网络不仅是大脑网络中连接最紧密最具弹性的的部分,还同时是存于意识网络中具有全局影响的部分——它是意识的真正核心。
论文题目:
How the brain transitions from conscious to subliminal perception
论文地址:
论文作者:
Francesca Arese Lucini, Gino Del Ferraro, Mariano Sigman, Hernan A. Makse
最近,以纽约城市学院 Francesca Arese Lucini 为首的四名科学家在arXiv发表了一篇预印本论文。研究者使用复杂网络中k-核渗透方法(k-core percolation)进行数学建模,观察了大脑由意识状态向无意识状态转变时功能网络的变化情况,发现意识状态下大脑功能网络有最多连接的核心部分(如视觉皮层和左额中回),与潜意识状态时保持活跃的区域重合,即意识与潜意识的功能网络内核是完全一致的。
该论文还解释了由意识到潜意识的转变过程:意识觉知状态因意识功能网络的外围k壳(k-shells)失活而丧失。这一发现论证了大脑功能网络最具鲁棒性的核心部分,对应于潜意识状态,而非之前一些理论所认为的意识状态,并对这些意识理论进行阐发。
意识状态向潜意识状态的转移
在这项研究中,作者首先通过对参考文献(Dehaene,2001)[1] 中 Dehaene等人于2001年中收集的功能磁共振成像数据进行了网络分析,研究了大脑从意识状态向潜意识转移的过程。
Dehaene 是法国神经学家,支持关于意识的全局空间理论(Global Workspace Theory),他曾在2001年与合作者们专门设计了两项实验,通过屏幕上闪现的诱导性单词,以分别监测人脑有意识和潜意识状态下的识字过程。
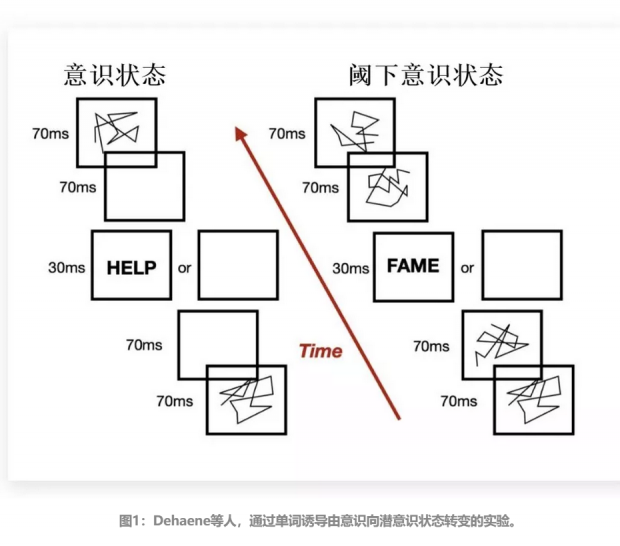
阈下(Subliminal)意识,指某种外在刺激未达到足够的强度仍在感觉阈限之下, 未引起大脑有意识觉知和注意力反应,但依然接收并处理了相关信息的一种潜意识过程。在 Dehaene 等的神经科学研究中,由阈下意识转向意识状态之前会有一系列神经活动的生理指标呈现。
如图1所示,左图描绘了引起有意识识别文字状态的一系列流程,其中四个字母的单词在电脑空白屏前后呈现,目的是让受试者有意识地感知这个单词,受试者每次都报告说看到了这个单词,这种fMRI状态信号被称为有意识的或无掩蔽的;而在右图描绘的实验中,单词被夹在干扰物之中,目的是让受试者不产生任何对单词的主动觉知。
这种用于区别受试者是否处于意识状态的方法,被称为主观报告法,是当前意识科学研究重要方法之一。最终实验结果表明,干扰图像确实起到了遮掩的作用,在其影响下受试者没有识别并意识到屏幕上的单词。这种fMRI状态信号称为掩蔽或潜意识的信号。
大脑活动立体激活图像
通过对以上对照实验受试者的大脑背景活动进行评估,将采集到的BOLD信号使用SPM99(UCL,1999)[2] 进行处理,然后再利用广泛采用的广义线性模型(Generalized Linear model)对fMRI进行时间序列分析(Smith,2004)[3],最终就得到了大脑的活动立体激活图像,如图2所示。
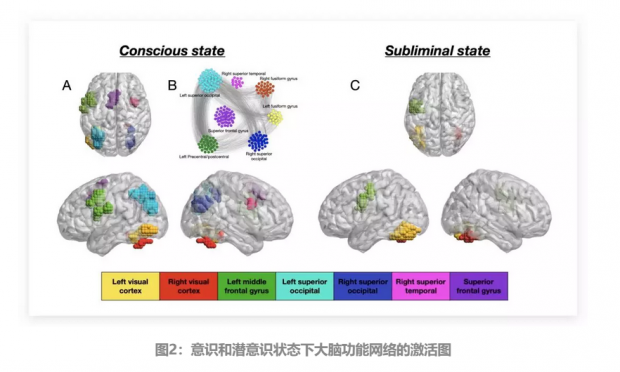
通过比较在图2a和图2c中标示出的意识状态和潜状态的激活情况,作者发现一些大脑区域在两种大脑状态中均参与活动。如视觉皮层(左视觉皮层-黄色、右视觉皮层-红色)和左额中回(绿色)——这些共有区域也是仅有的表征潜意识激活状态的大脑区域。
在意识状态被激活的实验中,功能性磁共振成像的激活会进一步扩散,并持续涉及到更多的其它集群。如图2a所示,它涉及到了左右枕上回(分别为浅蓝色和蓝色)、右额回(粉色)和感觉运动区(紫色)。
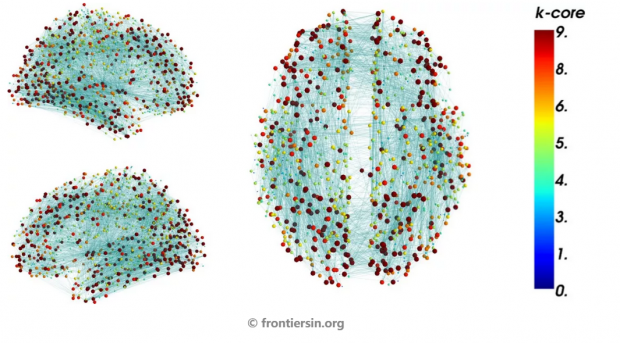
为了研究意识状态到潜意识状态转变的过程,作者根据意识状态实验中获得了扫描的fMRI激活图,并对每位受试者大脑分别构建了其功能网络状态的结构图,如图3。
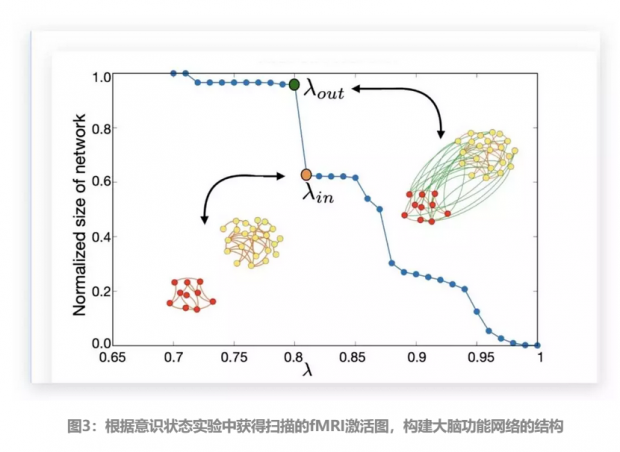
在为意识状态构建了大脑功能脑网络结构图后,作者使用复杂社会网络中k核渗透的方法,通过对属于不同k核的每一个结构中的节点进行了修剪操作,即k核分解(k-core decomposition),用以识别出两种意识状态下大脑结构网络中的共同模式和它们之间的差异。
什么是k核渗透?
k核(k-core)是数学中图论中的概念,最早在社会科学研究中被引入使用,原本用来描述社会网络的内聚性(Seidman,1983)[4],随后它被广泛应用于许多其它场合,如描述随机网络的鲁棒性(Dorogovtsev,2006)[5],社交网络中的病毒传播(Kitsak,2010)[6]以及大脑的大尺度结构(Hagmann,2008)[7]等。
对于给定的整个网络结构,k核是其中一个子图,作为子网络它由具有至少k个相互连接的所有临近节点组成。对某个子网络,k 的值有时被称为网络的核心度,它的大小与链接到子网络之外的实体节点数无关,只与子网络内互相连接的节点数大小有关。
对整个网络中的所有子网络,作为子图的k核其节点数k越大,说明网络中节点之间相互连接得越紧密。其中最大的 k 也被称为整个网络的最大核心度值 (k-values),例如你自己社交网络中的最大核心度就是你所有互为朋友圈的朋友的数目。
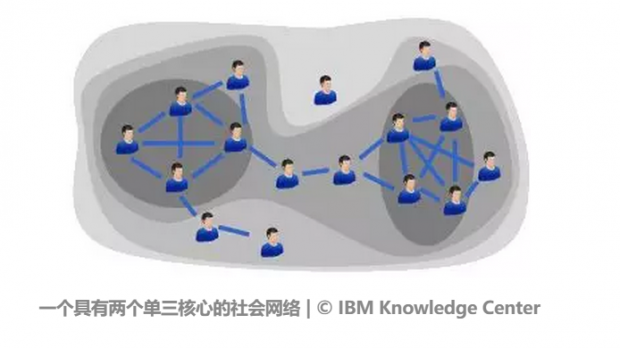
K-Core 与k-values的补充解释:
在上面网络中,包含了零核心、一核心、两核心和三核心的子网络嵌套集合,其中颜色最深的区域是左右两边的两个三核心子网络,网络中的节点都彼此与其他三个节点链接。而这两个三核心子网络,又包含在较浅颜色的一个二核心网络中。我们可以看到,零核心网络就相当于没有节点链接的全部孤立节点集合。对于给定核心度k的k核子网络的节点集合,总是0核包含1核,1核包含2核……k-1核包含k核,这是一个逐层嵌套结构。
对一个k核,因为总是包含在一个核心度为 k-1的子网络中,因此在这个子网络而不在k核的所有节点,就构成了k核的外围节点,它被称为k壳(k-shell)。同理,k核还包含在 k-1、k-2、……单核的子网络中,因为一个k核在整个网络中会具有多个k壳。
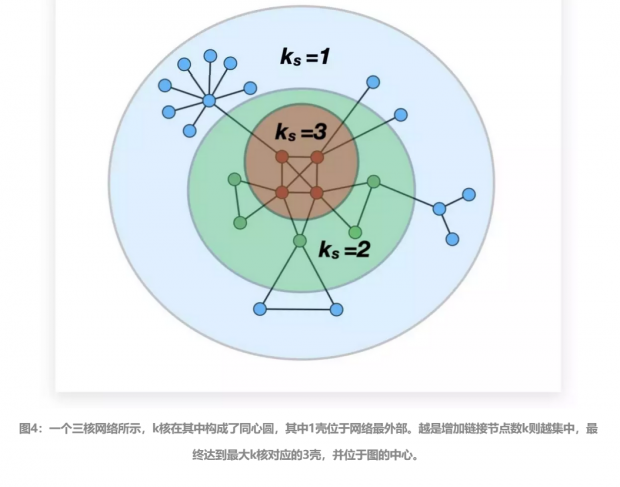
对具有不同k核子网络嵌套的整个网络来说,网络中最核心部分即对应于具有最大k核的子网络,它具有最大核心度值,称为Kmax核心,它网络的拓扑不变量(Dorogovtsev,2006)[5]。
对大脑功能网络的 k 核分解
对于给定的某个网络结构,要使用k核分解的方法,找到最大k核的子网络,可以通过迭代移除所有连接少于k的节点来获得:从k=1开始删除所有链接度小于k的节点,这些节点的移除会降低其临近节点的链接度。然后依次删除这些节点,并重复该过程,直到不能再删除更多节点。最后剩余的结构即是网络的最大k核心子网络。
“核心-外围结构”( core-periphery structure)
一旦我们按照上面的方法执行了k核的修剪过程,我们就可以计算出对每个给定的k,k核和k壳在其中每个受试者大脑活跃情况下所占的比率。
对于75个受试者意识状态全部大脑功能网络,论文作者都计算了k核和k壳的占用率,即在给定节点数k的情况下每个k核或k壳的在这些网络中的分布密度。当我们检查完每个k壳中的节点与大脑解剖对应的区域时,这些网络的一个有趣特性就显现出来了。
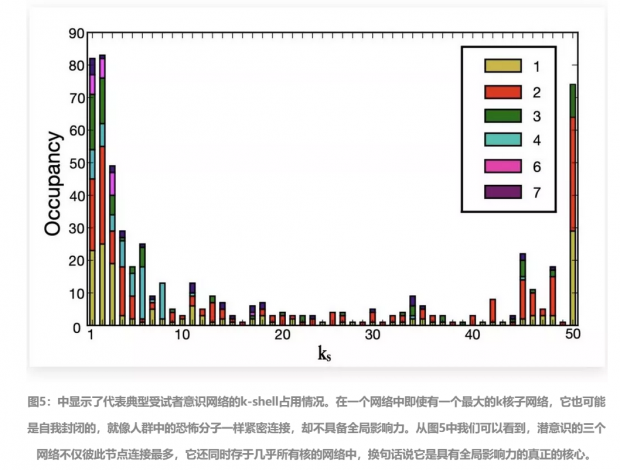
我们可以看到,非常有趣的是,大脑网络节点数量分布呈U形:对于非常小和非常高的k壳值,节点都显示出非常高的占用比,对于中间的k壳值,则显示出很低的占用比。
k最低的壳(1壳)由属于大脑中活跃的所有7个脑区簇的节点构成。相反最大k壳(如50壳),也即网络结构的中心内核,由仅属于3个脑区簇的节点构成,分别是左视觉皮层-黄色、右视觉皮层-红色以及左额叶中回-绿色——重要的是,它们是潜意识状态仅有的功能磁共振成像活跃的三个脑区。
这种U型分布形态在所有被分析的意识网络中都是一致的。
在参考文献(Burlesson-Lesser,2018)[8] 中将占用率呈U形分布的成因归于维持系统的稳定性:即在最低和最高k壳中的大量节点,使得网络能够同时对局部和全局随机攻击都具有很强的鲁棒性,从而使大脑成为随机扰动下的一个弹性系统(Resilient Systems)。
在生态系统和金融系统的网络中也能观察到同样的特征,事实上这是许多网络系统的一般特征,称为“核心-外围结构”( core-periphery structure)。(Corradino, 1990[9]; Krugman, 1996[10]; Borgatti & Everett, 2000[11]; Zhang et al. , 2015[12]; Verma et al. , 2016[13])
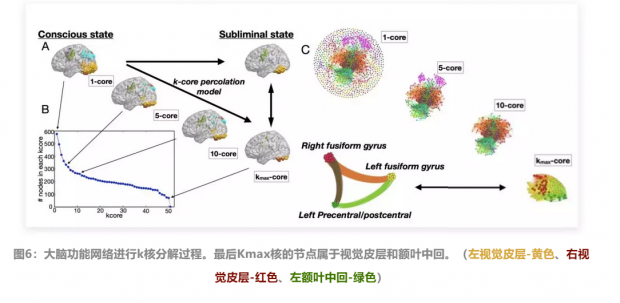
从上面图6,可以看到与图5显示相同的主题下大脑k核分解的过程:
图6a显示出,随着k核修剪,意识状态功能网络中的大脑的fMRI活动不断降低
图6b标识了每一个k核所占有的节点数。
图6c通过描绘大脑的网络连通性展示了k核分解的过程。
如前面图5所讨论那样,图6c最终显示出特定意识状态网络的Kmax核心由视觉核心和额叶中回构成,即平均潜意识状态仅有的功能磁共振成像激活的脑区集群。并且研究者通过随机网络验证实验,证明这样的结果并非由于随机效应。
最终论文表明,潜意识状态在无意识感知期间会持续保持活跃,并构成了意识状态的Kmax核心的很大一部分。
对当前意识理论的影响
Benjamin Libet 和 Dehaene 分别提出过两种关于无意识信息和有意识获取之间关系的重要理论。
意识谜题揭开前夜:
是源于信息处理过程?还是神经网络内在属性?
前者强调,通过对脑电图的数据分析可以表明,在任何有意志的行为产生之前几百毫秒里,所有外部刺激都首先是在大脑中无意识地处理好了的。根据这一理论,无意识信息是激起所有有意识行为的火花,而意识扮演的控制角色则对潜意识处理信息的执行者角色有调节作用(Libet,1985)[14]。
至于后者,Dehaene则已经证明在大脑中存在一个大规模多功能的脑区,它涉及顶叶和前额叶皮质的区域,该区域设置了一个临时的全局工作空间来绑定和共享信息(Dehaene & Naccache,2001[1];Dehaene,2014[15];Van Vugt,2018[16])。

意识的全局工作空间理论的支持者之一,巴黎法兰西公学院的认知科学家 Stanislas Dehaene
这种大脑先自下而上,再自上而下实现信息交互的框架被称为“点火”(ignition);如果外界输入的刺激没有足够强烈地激活脑内体素(voxels),那么大脑就不会将这些信息显示在意识中。
两种理论之间存在着共识,即虽然意识产生涉及大脑一系列大规模、非线性区域的激活,但这一过程开始于局部回路中,在为这个特定的过程编码信息后,意识才得以涌现。近日的这篇论文则提供了一个在双方共识基础之上的新观点。
这项研究表明,尽管会涉及大规模信息传播活动,无意识活动种子的内核却始终保持在功能网络壳结构的最核心处。这一发现让人想起David Mumford与同事(Lee, 1998)[17]提出的有关视觉的理论,该理论认为V1是大脑的高频功能核心,它起到暂时缓冲和保存最需要接收的字段信息的作用。
换句话说,意识激活的核心外壳可能并不是一组固定的神经元,而是在给定时间根据特定意识知觉的功能需求而变化状态的神经网络。
结论与思考
总体来说,该论文的研究成果与参考文献的预测是一致的(Morone,2019)[18],并能得出结论,意识到潜意识转变可以被解释为意识功能网络的外围k壳不断失活而丧失。最终残存的大脑区域,即最有弹性结构的区域Kmax核心,也就是那些构成潜意识状态的区域:视觉皮层和额叶中回。
这篇论文不仅揭示了意识到潜意识转变的本质,更强调了在系统神经科学中研究复杂网络结构的重要性。通过有效标记和建模,揭示意识和潜意识之间的相似性和差异性,从意识到潜意识觉知的转变过程,那么也许相反的,意识从潜意识浮现的探索之路也将会因为这种方法而走向下一阶段。
参考文献:
Dehaene, S, & Naccache, L. 2001. Towards a cognitive neuroscience of consciousness: basic evidence and a workspace framework. Cognition, 79, 1– 37.
UCL. 1999.
Smith, S M, Jenkinson, M, Woolrich, M W, Beckmann, C F, Behrens, T EJ, Johansen-Berg, H, Bannister, P R, De Luca, M, Drobnjak, I, Flitney, D E, et al. . 2004. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage, 23, S208–S219.
Seidman, S B. 1983. Network structure and minimum degree. Social Networks, 5, 269–287.
Dorogovtsev, S N, Goltsev, A V, & Mendes, J F F. 2006. K-core organization of complex networks. Physical Review Letters, 96, 040601.
Kitsak, M, Gallos, L K, Havlin, S, Liljeros, F, Muchnik, L, Stanley, H E, & Makse, H A. 2010. Identification of influential spreaders in complex networks. Nature Physics, 6, 888.
Hagmann, P, Cammoun, L, Gigandet, X, Meuli, R, Honey, C J, Wedeen, V J, & Sporns, O. 2008. Mapping the structural core of human cerebral cortex. PLoS Biology, 6, e159.
Burlesson-Lesser, K, Morone, F, & Makse, H A. 2018. K-core robustness in ecological and financial networks. submitted.
Corradino, C. 1990. Proximity structure in a captive colony of Japanese monkeys (Macaca fuscata fuscata): An application of multidimensional scaling. Primates, 31, 351–362.
Krugman, P R. 1996. The self-organizing economy. Blackwell Oxford.
Borgatti, S P, & Everett, M G. 2000. Models of core/periphery structures. Social Networks, 21, 375–395.
Zhang, X, Martin, T, & Newman, M E J. 2015. Identification of core-periphery structure in networks. Physical Review E, 91, 032803.
Verma, T, Russmann, F, Araujo, NAM, Nagler, J, & Herrmann, H J. 2016. ´ Emergence of core–peripheries in networks. Nature Communications, 7, 10441.
Libet, B. 1985. Unconscious cerebral initiative and the role of conscious will in voluntary action. Behavioral and Brain Sciences, 8, 529–539.
Dehaene, S. 2014. Consciousness and the brain: Deciphering how the brain codes our thoughts. Penguin.
Van Vugt, B, Dagnino, B, Vartak, D, Safaai, H, Panzeri, S, Dehaene, S, & Roelfsema, P R. 2018. The threshold for conscious report: Signal loss and response bias in visual and frontal cortex. Science, 360, 537–542.
Lee, TS. 1998. Mumford D, Romero R, and Lamme VA. The role of the primary visual cortex in higher level vision. Vision Res, 38.
Morone, F, Del Ferraro, G, & Makse, H A. 2019. The k-core as a predictor of structural collapse in mutualistic ecosystems. Nature Physics, 15, 95–102.
作者:十三维
编辑:王怡蔺
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}